
Blood
Alteration of microbial composition in the skin and blood in vasculitis
Sep
Abstract
Vasculitis is a systemic autoimmune disease characterized by leukocyte infiltration into blood vessels. Various microorganisms have been associated with the pathogenesis of vasculitis; however, the causal microbial agents and underlying mechanisms are not fully understood, possibly because of the technical limitations of pathogen detection. In the present study, we characterized the microbiome profile of patients with cutaneous vasculitis using comprehensive metagenome shotgun sequencing. We found that the abundance of the SEN virus was increased in the affected skin and serum of patients with vasculitis compared to healthy donors. In particular, the abundance of SEN virus reads was increased in the sera of patients with cutaneous arteritis. Among the bacteria identified, Corynebacteriales was the most differentially associated with vasculitis. Linear discriminant analysis effect size also indicated differences in the microbial taxa between patients with vasculitis and healthy donors. These findings demonstrate that vasculitis is associated with considerable alteration of the microbiome in the blood and skin and suggest a role for the infectious trigger in vasculitis.
Introduction
Vasculitis is a group of autoimmune diseases that cause various organ disorders, not only in the blood vessels but also in the central nerves, lungs, and kidneys1. Corticosteroids and immunosuppressive drugs are commonly used to treat vasculitis. However, the disease is intractable to the treatments in some cases; immunosuppression sometimes results in serious clinical complications, and a proper treatment for these conditions has not been established2. Since vasculitis is a relatively rare disease compared to other autoimmune diseases such as rheumatoid arthritis, it has not been studied sufficiently, and its etiology has not been fully elucidated3.
The association between vasculitis and various pathogens has been widely reported4. For example, immunoglobulin (Ig)A vasculitis is often triggered by a prior infection, such as an upper respiratory tract infection, caused by various microorganisms, including Streptococcus, Staphylococcus aureus, and varicella-zoster virus5. With regard to cutaneous small vessel vasculitis (CSVV), previously reported cause of the disease includes infections (20%), inflammatory conditions (15% to 20%), drug reactions (10% to 15%), or malignancies (5%), and the remaining half of CSVV cases are considered to be idiopathic6. We previously showed that a streptococcal antigen, the nephritis-associated plasmin receptor, is deposited in cutaneous vessels in IgA vasculitis, possibly inducing vascular inflammation7. Polyarteritis nodosa (PAN) is triggered by hepatitis B virus (HBV) infection8, and HBV on the vascular wall has been identified at the site of vasculitis9, suggesting that the pathogen may directly induce some types of vasculitis.
Studies on the association between vasculitis and pathogens have been conducted using animal models. For instance, injection of Candida albicans water-soluble fraction in mice induces inflammation of the aortic root and coronary arteries10,11,12,13, as well as in experimental models of vasculitis caused by viral, bacterial, fungal, and other pathogens4. Another study attempted to identify the causal microbe of the temporal arteries in patients with giant cell arteritis (GCA), and 16S ribosomal RNA gene sequencing revealed microbiome differences in temporal arteries between GCA and non-GCA. However, no single pathogen that causes GCA has been identified14.
With the advent of next-generation sequencing (NGS) technology, researchers have been able to obtain large amounts of data in a single round of operation15. NGS methods also enable the detection of microorganisms that are difficult to culture. Currently, most of these investigations use NGS by focusing on targeted amplicon sequencing (16S rRNA gene sequencing), or by sequencing the genetic material present within a sample directly using a PCR-independent approach (shotgun metagenomic sequencing)16. As shotgun metagenomic sequencing is PCR-independent, it comprehensively explores microorganisms and leads to the discovery of unknown novel pathogens that cannot be detected using targeted amplicon-based methods17. Indeed, a study comparing 16S rRNA gene sequencing with shotgun metagenomic sequencing for taxonomic characterization of gut microbiota showed that 16S rRNA gene sequencing detected only a fraction of the microbiota identified by shotgun sequencing18.
Among the various organs involved in vasculitis, the skin is the most privileged tissue because it is easily accessible for physical examinations and biopsies in comparison with other organs19. Thus, understanding the pathogenesis of vasculitis by identifying the causal microorganisms in the skin and blood could lead to a new therapeutic approach. In this study, we used NGS to analyze the microbial genome in skin and serum derived from patients with vasculitis.
Results
Viral profile of serum and skin tissue in vasculitis and HD (healthy donors)
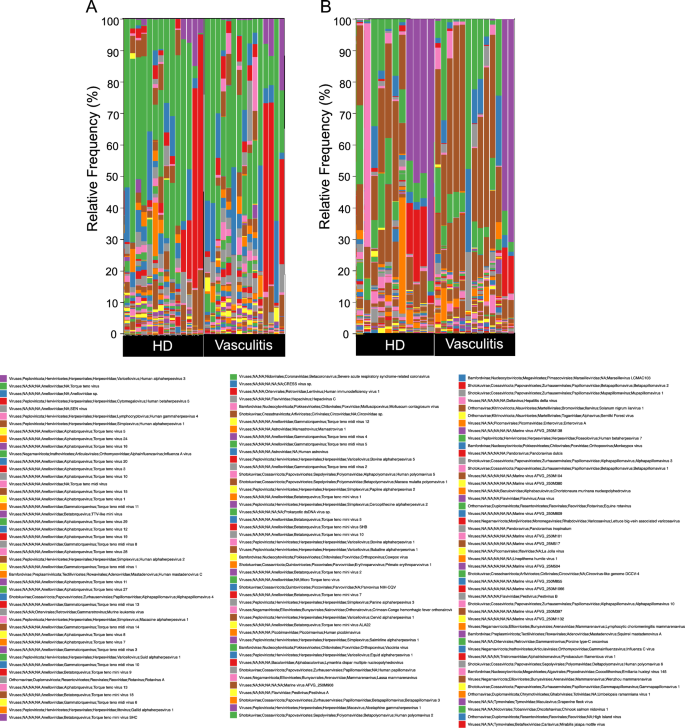
We first analyzed the viral composition in the serum and skin tissue using the Kraken2 software to determine if any specific species were enriched in the samples derived from patients with vasculitis. Interestingly, a large variety of viral species was detected in the serum (Fig. 1A), which is normally considered a sterile body fluid. The skin virome is also composed of many types of viruses (Fig. 1B), and the frequency of Human alphaherpesvirus 1 detected in the skin appeared to be higher than that in the serum (shown in brown). Among the viruses, the SEN virus (shown in gray) was more frequently detected in serum derived from patients with vasculitis (Fig. 2A), as well as in skin tissue derived from patients with vasculitis (Fig. 2B), than in HD (healthy donors).
Viral composition in patients with vasculitis and HD. Stacked graphs show the distribution of the relative frequency of viral species in serum (A) or skin tissue (B).

The proportion of SEN virus detected in patients with vasculitis and HD. Purple bar plot indicates relative frequency of SEN virus read counts in serum (A) or skin tissue (B).
The SEN virus has been linked to blood transfusion and is thought to be an important cause of post-transfusion hepatitis20. Although there is no reliable evidence that SEN virus infection leads to hepatitis, concurrent infections with the SEN virus, HBV, hepatitis C virus (HCV), or human immunodeficiency virus type 1 have been documented21. As HBV and HCV are associated with systemic vasculitis, we sought to investigate the possibility that the presence of the SEN virus influences the disease status of vasculitis.
SEN virus is abundantly detected in vasculitic skin tissue
We compared vasculitis patients with HD based on the abundance-normalized read counts (proportion of read counts) of the SEN virus. Although there was no significant difference in serum (Fig. 3A), the read counts of the SEN virus were significantly higher in skin tissue from patients with vasculitis than in HD (Fig. 3B). We also conducted qPCR to compare the SEN virus-positive rates in each serum sample. SEN virus was detected in 13 (76.5%) of the 17 patients with vasculitis and in 2 (40%) of the 5 HD (Table 1). The SEN virus has a tendency to be present in patients with vasculitis compared to those on HD. These results suggest that the SEN virus may be associated with skin symptoms in vasculitis. There was no remarkable correlation between the presence of the SEN virus and liver dysfunction in this patient cohort.

Abundance-normalized read counts (proportion of read counts) of SEN virus in patients with vasculitis and HD. (A) Serum read counts of SEN virus in patients with vasculitis and HD. (B) Tissue read counts of SEN virus in patients with vasculitis and HD. Values are mean ± SEM. *P < 0.05 vs. HD.
Serum viral reads of SEN virus based on patient’s characteristics
We further explored whether the reads of the SEN virus differed depending on the clinical features of the patients. The proportion of read counts for the SEN virus was significantly higher in patients with CA than in those on HD (Fig. 4A). However, there were no differences with regard to other clinical features, such as kidney involvement, use of corticosteroids, episode of preceding infection in IgA vasculitis, sex, or age in patients with vasculitis (Fig. 4B–F). Viral reads of the SEN virus in the skin tissue were not statistically different according to the clinical features of the patients with vasculitis (Fig. 5A–F). In HD, there was no statistical difference in the viral reads of the SEN virus by sex and age (Fig. S1, Supplementary Information).

Abundance-normalized serum read counts (proportion of read counts) of SEN virus by patient characteristics. (A) Type of vasculitis. (B) Kidney involvement. (C) Use of corticosteroids. (D) Episode of preceding infection in IgA vasculitis. (E) Sex. (F) Age. Values are mean ± SEM. *P < 0.05 vs. HD.

Abundance-normalized tissue read counts (proportion of read counts) of SEN virus by patient characteristics. (A) Type of vasculitis. (B) Kidney involvement. (C) Use of corticosteroids. (D) Episode of preceding infection in IgA vasculitis. (E) Sex. (F) Age. Values are mean ± SEM.
Bacterial composition in serum from patients with vasculitis and HD
IgA vasculitis is one of the most common types of vasculitis that present cutaneous symptoms. Although the mechanism in which the skin and kidney are involved in IgA vasculitis has not been clarified, numerous studies have linked IgA vasculitis with bacterial infection, particularly with β-hemolytic streptococci, as a possible trigger for the disease22. The majority of the patients enrolled in this study had IgA vasculitis. We therefore analyzed the bacterial composition in this cohort to determine whether bacteria were associated with the pathogenesis of vasculitis. The beta diversity of serum samples using PCA based on Euclidean distance showed a tendency toward a difference between vasculitis and HD (PERMANOVA: R2 = 0.06465, p = 0.0486) (Fig. 6A). There was no significant difference between the vasculitis and HD groups in terms of the Bray–Curtis distance (PERMANOVA: R2 = 0.05241, p = 0.05719) (Fig. 6B).

Circulating bacterial profile in patients with vasculitis and HD. Principal component analysis (PCoA) of serum from patients with vasculitis (black) compared to those from HD (red). P values less than 0.05 were considered as statistically significant.
We also analyzed serum bacterial taxa using LDA effect size (LEfSe) to detect differences between patients with vasculitis and HD. The bar plot shows linear discriminant analysis (LDA) scores of microbial taxa, with significant differences between the vasculitis and HD groups (Fig. 7A). The cladogram also indicated differences in microbial taxa between patients with vasculitis and those on HD (Fig. 7B). The bacterial read that was remarkably elevated in the vasculitis serum was from Corynebacteriales.

LDA effect size (LEfSe) analysis. (A) The bar plots represent the significantly differential taxa between vasculitis (green) or HD (red) (LDA score > 2). (B) Cladograms showing the taxa most differentially associated with vasculitis (green) or HD (red). Circle sizes in the cladogram plot are proportional to bacterial abundance. The circles represent, going from the inner to outer circle: phyla, genera, classes, order, and families. Yellow nodes represent species with no significant difference.
Discussion
The ability of leukocytes to enter tissues in response to immune stimuli is a central feature of the host defense23. On the other hand, excessively activated leukocytes, including lymphocytes, macrophages, and neutrophils, damage the vessels and aggravate inflammation12. While the etiology of vasculitis remains incompletely understood, an increasing number of animal and human studies have suggested that it is associated with a variety of pathogenic agents including bacteria, viruses, and fungi4. Previous studies have indicated that viruses relate to small-vessel vasculitis in many cases, whereas bacterial infections cause vasculitis in all sizes of vessels24.
In the present study, we utilized comprehensive metagenomic shotgun sequencing to demonstrate that the composition of viruses and bacteria in the serum and skin tissue is altered in vasculitis. Specifically, the frequency of the SEN virus detected in the skin tissue is elevated in vasculitis. In addition, the SEN virus is more likely to be present in the serum of patients with cutaneous arteritis than in those with IgA vasculitis.
The SEN virus is a blood-borne single-stranded circular DNA virus discovered in 199921. The prevalence of the SEN virus has been reported to be 10% in Japan25, with a higher incidence among patients with liver diseases in comparison with the general population. Among the nine different genetic variants of the SEN virus (A to I), the most predominant genotypes are SEN virus D and H, which are typically found in patients with non-A-E hepatitis26. HBV is associated with cryoglobulinemic small-vessel vasculitis and polyarteritis nodosa (PAN), and the relationship between PAN and HBV has been often reported4. Approximately 5% of patients with HCV-associated mixed cryoglobulinemia develop cryoglobulinemic vasculitis, as a consequence of the deposition of immune complex (IC) in small vessels. Although we were not able to histologically verify the presence of the SEN virus in the skin vessels owing to a lack of available SEN virus-specific antibodies, it is possible that direct injury of the vessel wall or IC deposition in the vessel wall induces vascular inflammation, similar to other viruses. In addition, IgA vasculitis might be a more heterogeneous condition than CA, based on the finding that the frequency of the SEN virus was higher in serum samples from CA, but not in those from IgA, compared to HD.
In the present study, we examined the bacterial composition of patients with vasculitis on HD. No remarkable changes in bacterial diversity were detected in the sera of patients with vasculitis and HD. However, the abundance of bacterial species was considerably different between the sera derived from patients with vasculitis and HD. Among the bacteria identified in the serum, Corynebacteriales were the most differentially associated with vasculitis. The genus Corynebacterium is a taxon of coryneform gram-positive bacilli or coccobacilli that colonize the mucous membranes and skin in humans27. Corynebacterium is usually regarded as a clinically non-significant contaminant in cultures28, but is sometimes reported in the context of autoimmune diseases. A previous study suggested that Corynebacterium 1 in the gut was associated with disease duration, and serum IL-1β levels in patients with vitiligo29. Other studies have implicated that the presence of Corynebacterium in the nasopharyngeal microbiome30 and peripheral blood leukocytes31 relates to Kawasaki disease. In the present study, although no bacteria already known to be associated with IgA vasculitis were detected, such as Staphylococcus and Streptococcus species, Corynebacterium species may serve as novel pathogenic candidate microbes for vasculitis.
Some limitations in this study need to be acknowledged. First, we detected numerous microorganisms using metagenome sequences; however, contamination in the blood or skin should always be considered. Although we removed the epidermis before gene extraction to exclude resident skin flora, the presence of Corynebacterium in the cutaneous vessels should be further verified as Corynebacterium is one of the major skin-resident bacteria32. Second, the small sample size and limited types of vasculitis may have affected the results. Third, we did not assess whether the microorganisms were truly pathogenic or were present only in the blood or skin tissue. Further studies using animal models should provide important insights into the pathogenic role of microbiome alterations in vasculitis.
In conclusion, we found specific alterations in the microbiome in cutaneous vasculitis and identified new candidates for the pathogenic microbe using metagenomic shotgun analysis. These alterations of microbial composition may act as environmental triggers or potentially aggravating factors for vasculitis. Further studies are required to verify the importance of these pathogens on the initiation and progression of vasculitis.
Materials and methods
Study population
Blood and skin tissue samples were collected from 18 patients with vasculitis who visited our division between 2020 and 2022 and 14 HD. Participants were divided into patients diagnosed with cutaneous arthritis (CA; n = 4), IgA vasculitis (n = 12), microscopic polyangiitis (MPA; n = 1), and cryoglobulinemic vasculitis (n = 1). The authors had access to information that could identify individual participants during or after data collection.
Clinical information is shown in Table 2. There were no significant differences between the patients with vasculitis and those on HD with regard to age or sex. One patient with IgA vasculitis had a history of HBV virus infection. Skin biopsies were taken from the active cutaneous vasculitis lesions in lower legs, except one case of IgA vasculitis taken from the knee. The experimental protocol was approved by the Ethics Committee of Tokyo Women’s Medical University (approval number: 5700) and all subjects provided written informed consent according to the principles of the Declaration of Helsinki.
DNA and RNA extraction from skin tissue and blood samples
Skin tissue samples were cut into small pieces, vortexed with Phosphate-buffered saline (PBS), and centrifuged to collect the supernatant. The QIAamp MinElute Virus Spin Kit (Qiagen, Hilden, Germany) was used for DNA and RNA extraction from supernatant fractions and blood samples. The quantities of the collected DNA and RNA were measured using a Qubit fluorometer (Thermo Fisher Scientific).
Metagenomic shotgun sequencing
The extracted RNA was converted into double-stranded copy (c)DNA using the ProtoScript II First Strand cDNA Synthesis Kit (New England Biolabs, Ipswich, MA, USA), NEBNext Ultra II Non-Directional RNA Second Strand Synthesis Module (New England Biolabs), and Random Primer 6 (random hexanucleotides; New England Biolabs). The obtained double-stranded (ds)DNA and DNA samples were subsequently converted to Illumina TruSeq-compatible libraries using Twist Library Preparation Enzymatic Fragmentation Kit (Twist Biosciences, CA, USA) and Twist Universal Adapter System-TruSeq Compatible (Twist Biosciences). We performed a single hybridization capture for each sample and enrichment using the Twist Comprehensive Viral Research Panel (Twist Biosciences). All libraries were converted into libraries for the DNBSEQ using the MGIEasy Universal Library Conversion Kit (App-A). Sequencing was performed using the DNBSEQ-G400RS High-throughput Sequencing Kit (MGI Tech, Tokyo, Japan) in 100-base paired-end mode. Each read was subjected to KRAKEN2 analysis against the PlusPFP database containing archaea, bacteria, viral, plasmid, human, UniVec_core, protozoa, fungi, and plant33,34.
Extraction of serum DNA and quantitative real-time polymerase chain reaction
DNA extraction from 200 μl of serum was carried out with the Plasma/Serum Cell-free Circulating DNA Purification Mini Kit (Norgen, Thorold, Canada) as per protocol. Quantitative polymerase chain reactions (qPCR) in the presence of SYBR Green (PowerUp SYBR Green Master Mix, Applied biosystems, MA, USA) were performed with a QuantStudio 3 (Thermo Fisher Scientific, MA, USA) using the following primers: AI, 5’- TWCYCMAACGACCAGCTAGACCT—3’ (forward) and 5’- GTTTGTGGTGAGGAGAACGGA—3’ (reverse)35. Cycle threshold of less than 35 defined a positive result.
Statistical analysis
Data were analyzed using Prism 9 (GraphPad Software), and results were expressed as the mean ± SEM. Values between the two groups were compared using an unpaired 2-tailed Student’s t-test. P values for multiple groups were calculated using ordinary one-way ANOVA with Dunnett’s post hoc test. P value for the contingency table was calculated using Fisher’s exact test. A value of P < 0.05 defined the presence of a statistically significant difference.
Data availability
The data that support the findings of this study are available from the corresponding author, [CM], upon reasonable request.
References
-
Jennette, J. C. et al. 2012 revised International Chapel Hill consensus conference nomenclature of vasculitides. Arthritis Rheum. 65, 1–11. https://doi.org/10.1002/art.37715 (2013).
Google Scholar
-
Miyabe, Y., Miyabe, C., Iwai, Y. & Luster, A. D. Targeting the chemokine system in rheumatoid arthritis and vasculitis. Jma J. 3, 182–192. https://doi.org/10.31662/jmaj.2020-0019 (2020).
Google Scholar
-
Watts, R. A. & Robson, J. Introduction, epidemiology and classification of vasculitis. Best Pract. Res. Clin. Rheumatol. 32, 3–20. https://doi.org/10.1016/j.berh.2018.10.003 (2018).
Google Scholar
-
Miyabe, C., Miyabe, Y., Miyata, R. & Ishiguro, N. Pathogens in vasculitis: Is it really idiopathic?. Jma J. 4, 216–224. https://doi.org/10.31662/jmaj.2021-0021 (2021).
Google Scholar
-
Sugino, H., Sawada, Y. & Nakamura, M. IgA vasculitis: Etiology, treatment, biomarkers and epigenetic changes. Int. J. Mol. Sci. https://doi.org/10.3390/ijms22147538 (2021).
Google Scholar
-
Shavit, E., Alavi, A. & Sibbald, R. G. Vasculitis-what do we have to know? A review of literature. Int. J. Low Extrem. Wounds 17, 218–226. https://doi.org/10.1177/1534734618804982 (2018).
Google Scholar
-
Miyabe, C., Oda, T., Miyata, R., Miyabe, Y. & Ishiguro, N. Nephritis-associated plasmin receptor in the cutaneous vessels in IgA vasculitis. J. Dermatol. 50, 102–103. https://doi.org/10.1111/1346-8138.16574 (2023).
Google Scholar
-
Ishiguro, N. & Kawashima, M. Cutaneous polyarteritis nodosa: A report of 16 cases with clinical and histopathological analysis and a review of the published work. J. Dermatol. 37, 85–93. https://doi.org/10.1111/j.1346-8138.2009.00752.x (2010).
Google Scholar
-
Trepo, C. & Guillevin, L. Polyarteritis nodosa and extrahepatic manifestations of HBV infection: the case against autoimmune intervention in pathogenesis. J. Autoimmun. 16, 269–274. https://doi.org/10.1006/jaut.2000.0502 (2001).
Google Scholar
-
Miyabe, C. et al. Am 80, a retinoic acid receptor agonist, ameliorates murine vasculitis through the suppression of neutrophil migration and activation. Arthritis Rheum. 65, 503–512. https://doi.org/10.1002/art.37784 (2013).
Google Scholar
-
Miyabe, C. et al. A sphingosine 1-phosphate receptor agonist ameliorates animal model of vasculitis. Inflamm. Res. 66, 335–340. https://doi.org/10.1007/s00011-016-1018-y (2017).
Google Scholar
-
Miyabe, C. et al. Abrogation of lysophosphatidic acid receptor 1 ameliorates murine vasculitis. Arthritis Res. Ther. 21, 191. https://doi.org/10.1186/s13075-019-1973-0 (2019).
Google Scholar
-
Miyabe, C. et al. Dectin-2-induced CCL2 production in tissue-resident macrophages ignites cardiac arteritis. J. Clin. Invest. 129, 3610–3624. https://doi.org/10.1172/jci123778 (2019).
Google Scholar
-
Hoffman, G. S. et al. The microbiome of temporal arteries. Pathog. Immun. 4, 21–38. https://doi.org/10.20411/pai.v4i1.270 (2019).
Google Scholar
-
Motooka, D., Nakamura, S., Hagiwara, K. & Nakaya, T. Viral detection by high-throughput sequencing. Methods Mol. Biol. 1236, 125–134. https://doi.org/10.1007/978-1-4939-1743-3_11 (2015).
Google Scholar
-
Boers, S. A., Jansen, R. & Hays, J. P. Understanding and overcoming the pitfalls and biases of next-generation sequencing (NGS) methods for use in the routine clinical microbiological diagnostic laboratory. Eur. J. Clin. Microbiol. Infect. Dis. 38, 1059–1070. https://doi.org/10.1007/s10096-019-03520-3 (2019).
Google Scholar
-
Nakamura, S., Nakaya, T. & Iida, T. Metagenomic analysis of bacterial infections by means of high-throughput DNA sequencing. Exp. Biol. Med. (Maywood) 236, 968–971. https://doi.org/10.1258/ebm.2011.010378 (2011).
Google Scholar
-
Durazzi, F. et al. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 11, 3030. https://doi.org/10.1038/s41598-021-82726-y (2021).
Google Scholar
-
Miyabe, C. et al. Immune checkpoint molecule expression is altered in the skin and peripheral blood in vasculitis. Sci. Rep. 11, 20019. https://doi.org/10.1038/s41598-021-99558-5 (2021).
Google Scholar
-
Almaqati, T. et al. Molecular epidemiology of SEN virus among blood donors and renal dialysis patients. Acta Biomed. 93, e2022237. https://doi.org/10.23750/abm.v93i5.13005 (2022).
Google Scholar
-
Akiba, J., Umemura, T., Alter, H. J., Kojiro, M. & Tabor, E. SEN virus: Epidemiology and characteristics of a transfusion-transmitted virus. Transfusion 45, 1084–1088. https://doi.org/10.1111/j.1537-2995.2004.00209.x (2005).
Google Scholar
-
Brogan, P. & Eleftheriou, D. Vasculitis update: Pathogenesis and biomarkers. Pediatr. Nephrol. 33, 187–198. https://doi.org/10.1007/s00467-017-3597-4 (2018).
Google Scholar
-
Spertini, O., Kansas, G. S., Munro, J. M., Griffin, J. D. & Tedder, T. F. Regulation of leukocyte migration by activation of the leukocyte adhesion molecule-1 (LAM-1) selectin. Nature 349, 691–694. https://doi.org/10.1038/349691a0 (1991).
Google Scholar
-
Somer, T. & Finegold, S. M. Vasculitides associated with infections, immunization, and antimicrobial drugs. Clin. Infect. Dis. 20, 1010–1036. https://doi.org/10.1093/clinids/20.4.1010 (1995).
Google Scholar
-
Shibata, M. et al. The presence of a newly identified infectious agent (SEN virus) in patients with liver diseases and in blood donors in Japan. J. Infect. Dis. 184, 400–404. https://doi.org/10.1086/322050 (2001).
Google Scholar
-
Pirouzi, A., Bahmani, M., Feizabadi, M. M. & Afkari, R. Molecular characterization of Torque teno virus and SEN virus co-infection with HIV in patients from Southern Iran. Rev. Soc. Bras. Med. Trop. 47, 275–279. https://doi.org/10.1590/0037-8682-0073-2014 (2014).
Google Scholar
-
Tauch, A., Fernández-Natal, I. & Soriano, F. A microbiological and clinical review on Corynebacterium kroppenstedtii. Int. J. Infect. Dis. 48, 33–39. https://doi.org/10.1016/j.ijid.2016.04.023 (2016).
Google Scholar
-
Yamamuro, R., Hosokawa, N., Otsuka, Y. & Osawa, R. Clinical characteristics of corynebacterium bacteremia caused by different species, Japan, 2014–2020. Emerg. Infect. Dis. 27, 2981–2987. https://doi.org/10.3201/eid2712.210473 (2021).
Google Scholar
-
Ni, Q. et al. Gut microbial dysbiosis and plasma metabolic profile in individuals with vitiligo. Front. Microbiol. 11, 592248. https://doi.org/10.3389/fmicb.2020.592248 (2020).
Google Scholar
-
Sánchez-Manubens, J. et al. Characterization of the nasopharyngeal microbiome in patients with Kawasaki disease. An. Pediatr. (Engl. Ed.) 97, 300–309. https://doi.org/10.1016/j.anpede.2022.08.001 (2022).
Google Scholar
-
Shibata, M., Ezaki, T., Hori, M., Nagashima, M. & Morishima, T. Isolation of a Kawasaki disease-associated bacterial sequence from peripheral blood leukocytes. Pediatr. Int. 41, 467–473. https://doi.org/10.1046/j.1442-200x.1999.01115.x (1999).
Google Scholar
-
Chen, Y. E., Fischbach, M. A. & Belkaid, Y. Skin microbiota-host interactions. Nature 553, 427–436. https://doi.org/10.1038/nature25177 (2018).
Google Scholar
-
Lu, J. et al. Metagenome analysis using the Kraken software suite. Nat. Protoc. 17, 2815–2839. https://doi.org/10.1038/s41596-022-00738-y (2022).
Google Scholar
-
Wood, D. E., Lu, J. & Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 20, 257. https://doi.org/10.1186/s13059-019-1891-0 (2019).
Google Scholar
-
Kojima, H., Kaita, K. D., Zhang, M., Giulivi, A. & Minuk, G. Y. Genomic analysis of a recently identified virus (SEN virus) and genotypes D and H by polymerase chain reaction. Antiviral Res. 60, 27–33. https://doi.org/10.1016/s0166-3542(03)00133-5 (2003).
Google Scholar
Acknowledgements
We thank Ms. Mai Onuki for technical assistance.
Funding
This work was supported by Takeda Medical Research Foundation Research Grant, GSK Japan Research Grant 2021, and Medical Research Encouragement Prize of The Japan Medical Association.
Author information
Authors and Affiliations
Contributions
R.M. and C.M. designed, collected patient samples, analyzed and interpreted data, and wrote the manuscript. H.O., D.M., and S.N. performed Metagenomic shotgun sequencing. Y.M., Y.T., Y.F., and K.Y. supervised the project. N.I. provided overall project supervision and contributed to the design of the experiments. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Figure S1.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Reprints and Permissions
About this article
Cite this article
Miyata, R., Miyabe, C., Oki, H. et al. Alteration of microbial composition in the skin and blood in vasculitis.
Sci Rep 13, 15317 (2023). https://doi.org/10.1038/s41598-023-42307-7
-
Received: 17 May 2023
-
Accepted: 07 September 2023
-
Published: 15 September 2023
-
DOI: https://doi.org/10.1038/s41598-023-42307-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.