
Cardiovascular
The therapeutic value of bifidobacteria in cardiovascular disease
Oct
Abstract
There has been an increase in cardiovascular morbidity and mortality over the past few decades, making cardiovascular disease (CVD) the leading cause of death worldwide. However, the pathogenesis of CVD is multi-factorial, complex, and not fully understood. The gut microbiome has long been recognized to play a critical role in maintaining the physiological and metabolic health of the host. Recent scientific advances have provided evidence that alterations in the gut microbiome and its metabolites have a profound influence on the development and progression of CVD. Among the trillions of microorganisms in the gut, bifidobacteria, which, interestingly, were found through the literature to play a key role not only in regulating gut microbiota function and metabolism, but also in reducing classical risk factors for CVD (e.g., obesity, hyperlipidemia, diabetes) by suppressing oxidative stress, improving immunomodulation, and correcting lipid, glucose, and cholesterol metabolism. This review explores the direct and indirect effects of bifidobacteria on the development of CVD and highlights its potential therapeutic value in hypertension, atherosclerosis, myocardial infarction, and heart failure. By describing the key role of Bifidobacterium in the link between gut microbiology and CVD, we aim to provide a theoretical basis for improving the subsequent clinical applications of Bifidobacterium and for the development of Bifidobacterium nutritional products.
Introduction
Cardiovascular disease (CVD) is the leading cause of death worldwide, accounting for ~30% of global mortality1. According to the World Health Organization (WHO), the global death toll from CVD was 17.9 million in 20212. Physiological factors such as age, gender, and genetics as well as poor daily behavior are major CVD factors3. In addition, pathological states such as inflammation and oxidative stress and underlying diseases such as obesity, hyperlipidaemia (HLP), as well as diabetes have been proven to cause or exacerbate CVD3,4. However, a growing body of data has shown a correlation between intestinal microbiota and CVD, and that such microbiota hold great promise as an emerging therapeutic tool5. It has been reported that homeostasis of the intestinal microbiota, its physiological functions, and metabolites modulate human physiopathological processes and influence the developmental of CVD6.
Bifidobacterium belongs to the phylum Actinobacteria, a genus of high G + C Gram-positive, non-motile, rod-shaped, strictly anaerobic bacteria that are widely found in the intestinal tract and other luminal tissues of humans and animals7,8,9. As one of the first microorganisms to colonize the human gut and to accompany humans throughout their lives, bifidobacteria have a wide range of benefits for human health8. Bifidobacteria are one of the most important bacterial groups in the neonatal gut which promote proper development of physiological functions in neonates. In the early stages of life, bifidobacteria comprise up to 90% of the infant gut microbiota8,10. In the gut of adults, bifidobacteria make up 2–14% of the total gut microbial community; in the elderly, this number shrinks once again11. As probiotics, bifidobacteria are closely associated with human health and are best known for their role in promoting the health of the immune, digestive, and metabolic systems8.
Recent advances in research have shown that in the gut, which is rich in more than 1000 microorganisms, bifidobacteria show a significant potential for improving cardiovascular disease12. Interestingly, the abundance of bifidobacteria tended to decrease when cardiovascular events occurred, and conversely, their abundance was upregulated when CVD was ameliorated13,14,15. Emerging scientific research evidence suggested that bifidobacteria not only has a mitigating effect on inflammation, oxidative stress, but also improves intestinal barrier function as well as regulates intestinal microbial metabolites14,16,17,18. Furthermore, researchers have shown that obesity, HLP, and type 2 diabetes mellitus (T2DM), which are serious risk factors for CVD, could indirectly be reduced by bifidobacteria19,20,21. This review has mainly discussed the direct and indirect mechanisms of bifidobacteria in inhibiting the development of CVD, including suppression of oxidative stress, immunomodulation, and correction of lipid, glucose, and cholesterol metabolism to reduce classical risk factors for CVD. In addition, current advances in the use of bifidobacteria in hypertension (HP), atherosclerosis (AS), myocardial infarction (MI), and heart failure (HF) are discussed with an outlook on the future use of bifidobacteria as a cutting-edge therapeutic strategy for the prevention or treatment of CVD.
Direct improvement of cardiovascular disease
Antioxidant properties
Oxidative stress exists in various CVDs, such as AS, HP, myocardial ischemia-reperfusion (I/R) injury and HF, and strongly plays a role in its development or exacerbation22. The oxidative stress process occurs when there is production of excess reactive oxygen species (ROS), which include superoxide radicals, peroxyl radicals, hydroxyl radicals and hydrogen peroxide23. In AS, oxidative stress first causes endothelial dysfunction, thus increasing vascular permeability24. The oxidative stress then modifies the low-density lipoprotein (LDL) recruited to the artery walls into oxidized low-density lipoprotein (ox-LDL), which forms the basis for foam cells24. Finally, the increased ROS exacerbates plaque development24. On the other hand, ROS acts on vascular endothelial cell phosphorylation pathways and gene expression factors in HP to cause abnormal vascular tone. Myocardial I/R manifests as restoration of blood flow to hypoxic organs, a process that leads to excessive ROS production25. In addition, all phases of HF are accompanied by an excessive production of ROS24. This demonstrates that oxidative stress on the heart aggravates the prevailing CVD.
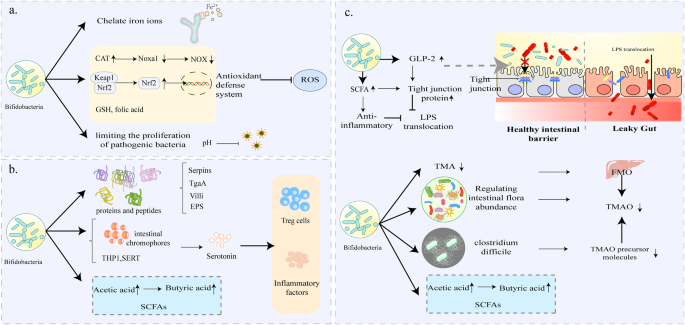
Many studies have demonstrated that bifidobacteria can improve CVD due to its high antioxidant activity, for example, Bifidobacterium longum subsp. longum (B. longum) CCFM752 prevented HP and aortic lesions in rats26, and its culture supernatant prevented Ang II-induced increases in O2- and H2O2 in smooth muscle A7R5 cells of rat thoracic aorta16. Previously, it was observed that an increase in the number of bifidobacteria in the intestine was resulted in reduced cardiac risk ratios and atherosclerotic indices in broilers27. Numerous data on the mechanism of antioxidant action of bifidobacteria is available. First, iron-binding sites were identified in various bifidobacteria species, including B. longum NCC2705, which suggests its ability to chelate iron ions, thus inhibiting its catalytic effect on oxidation reactions (Fig. 1a)23. Catalase (CAT), which degrades hydrogen peroxide, has been widely shown to be a critical enzyme in the antioxidant defense system. Previous data showed that CAT activity was enhanced and NADPH oxidase (NOX) activity was inhibited in A7R5 cells preincubated with culture supernatant of B. longum CCFM752 (Fig. 1a)16. Interestingly, the enhancement of CAT activity was not due to changes at the transcriptional level but to the upregulation of related proteins at the translational level, as cellular transcriptome sequencing revealed that the 60S ribosomal protein L7a (Rpl7a) was upregulated and its expression was positively correlated with intracellular CAT activity. As for the suppression of NOX activity, it may be due to the decreased expression of Noxa1. Other studies have demonstrated that Nrf2 – Keap1 – ARE is a pivotal pathway in the antioxidant defense system (Fig. 1a). Upon exposure to oxidative stress, the active cysteine residues of Keap1 are modified, resulting in inactivation of Keap1. This allows Nrf2 to bind Keap1 to form a complex that is ubiquitinated. Next, the accumulated Nrf2 acts as a master regulatory transcription factor, activating phase II detoxification enzymes, drug transporters, and antioxidant enzymes, thereby exerting intracellular antioxidant defense28. Previous report have shown that bifidobacteria attenuate oxidative stress by promoting the dissociation of Nrf2 from Keap1 and facilitating the translocation of Nrf2 to the nucleus. (Fig. 1a)29. Multiple explanations have been proposed to account for the mechanism by which bifidobacteria induce the expression of Nrf2. The mechanism underlying the ability of Bifidobacterium to induce Nrf2 expression remained unclear until researchers directed their attention toward certain intestinal metabolites. The first is indole-3-lactic acid (ILA), a major catabolic metabolite of tryptophan produced by more than 50 Bifidobacterium species30, and in 2020 researchers found that ILA increased the mRNA expression of GPX2, SOD2, and NQO1, the target activator genes of Nrf231. Second, an intestinal metabolite called sedanolide has recently been reported as a possible key effector in the activation of the Nrf2 pathway by bifidobacteria. Because the increase in sedanolide abundance in mice intestine, following administration of B. longum R0175, resulted in a significant upregulation of Nrf2 expression and Nrf2 pathway-related genes (NQO1, HO1)32. In addition, bifidobacteria metabolites, such as glutathione (GSH) and folic acid, also exert antioxidant benefits by maintaining the redox state of cells and increasing the antioxidant property of lipoproteins, respectively (Fig. 1a)33. It has also been proposed that the acid-producing effect of bifidobacteria could help maintain a low intestinal pH, limiting the proliferation of pathogenic bacteria, thus contributing to anti-oxidative stress (Fig. 1a)33. In summary, the anti-oxidative stress activity of bifidobacteria is realized by trapping metal ions, activating the antioxidant enzyme system and producing antioxidant metabolites which regulate intestinal microbiota.
a Bifidobacterium may function as an iron ion chelator to inhibit oxidation and oxidation reactions. It can activate the key genes in the antioxidant defense system, produce glutamic acid (GSH) to maintain the cellular redox state, and inhibit the production of pathogenic bacteria through the acidic environment. b Bifidobacterium has special molecular structures such as Serpins, villi, TgaA, and EPS that allow it to participate in specific immunomodulatory processes and cause anti-inflammatory activity. Its metabolite acetate can also stimulate the production of butyrate, which has anti-inflammatory effects, by cross-feeding as a substrate. c Bifidobacterium can act on intestinal epithelial cell junctions to maintain the integrity of the intestinal barrier and inhibit LPS translocation; it can also protect the intestinal barrier from damage by controlling the production of TMAO, SCFAs and maintaining the abundance of intestinal microbial species. (The figure does not contain any third-party material, the figure and each of the elements in the figure were created by the authors).
Immunoregulatory properties
Previous data have shown that inflammation is critical in the development of CVD, especially AS. Activation of various inflammatory signaling pathways and increased production of pro-inflammatory cytokines depict intensification of inflammatory response, which directly relates to CVD such as AS, I/R, and MI34. Furthermore, the inflammatory response could influence the prognosis of some CVD. For instance, a previous study demonstrated the association between the intensity of the inflammatory response and ventricular remodeling after MI35.
The immunoregulatory properties of bifidobacteria are well known and thus have long been used to treat several inflammatory conditions, such as allergies and necrotizing enterocolitis in premature infants. The accumulating knowledge about intestinal microbial functions has allowed researchers to focus on the relationship between bifidobacteria and CVD36. Liang et al.14 reported that Bifidobacterium animalis subsp. lactis (B. lactis) F1-7 enhanced the anti-inflammatory effect of krill oil, thereby alleviating inflammatory response in atherosclerotic mice more strongly compared to administration of krill oil alone. B. lactis has been reported in a randomized trial to significantly reduce the pro-inflammatory cytokines TNF-α and IL-6 in blood samples from patients with metabolic syndrome, which contributes to the reduction in cardiovascular risks in patients with metabolic syndrome15. Several specific molecular structures in bifidobacteria, such as secreted immunomodulatory proteins and peptides (Serpins, villi, peptidoglycan hydrolase TgaA) as well as extracellular polysaccharides (EPS), contribute to its anti-inflammatory properties (Fig. 1a)37. These specialized structures drive specific immunomodulatory processes by interacting with intestinal microbes or host cells. Specifically, production of serpin protects the bifidobacteria from adverse effects of host-derived proteases through anti-inflammatory activities38. Some villi (sorting enzyme-dependent hairs) could induce high levels of local inflammatory factors while inhibiting systemic inflammatory response, thus facilitating initial crosstalk between bifidobacteria and host cells38. Peptidoglycan hydrolase TgaA has been shown to induce IL-2 production in key cytokines in T cells39. EPS affects the differentiation of T cells into T helper cells thus regulating the levels of pro-inflammatory and anti-inflammatory cytokines. In addition, it was shown that bifidobacteria could trigger intestinal metabolite cross-feeding mechanisms and maintain intestinal homeostasis, which contributes to the maintenance of the intestinal immune system (Fig. 1a)37,38. Among the intestinal metabolites, butyrate is the potent anti-inflammatory agent that maintains the immune system and is involved in immune signaling by stimulating anti-inflammatory factors (TNF-β, IL-10) and inducing T-cell differentiation. Although bifidobacteria do not directly produce butyrate, the produced acetate can be used as a fermentation substrate for the indirect production of butyrate through cross-feeding interaction40. Strains of Bifidobacterium species also participate in tryptophan metabolism in the gut, while its metabolite indole-3-carbaldehyde (I3C) directly binds to AhR ligands involved in immune signaling and inhibits abnormal immune responses41,42. Although serotonin is well known as a central nervous system neurotransmitter, there is growing evidence that peripheral serotonin affects cardiovascular disease by influencing immune cell reactivity43. Previous studies have shown that bifidobacteria reduces serotonin levels by modulating key components of the serotonergic system in different host tissues, the number of intestinal chromophores and gene expression during serotonin synthesis and reuptake (THP1, SERT)44,45,46,47. In fact, these are far from allowing us to fully understand the role of bifidobacteria on the human immune system, as most of the current research is still at the cellular or animal level. Given the intricate nature of intestinal microorganisms, deciphering the immunomodulatory mechanisms of bifidobacteria within the gut presents a formidable task. It’s also important to acknowledge the potential influence, and even reversal, of bifidobacteria’s immunomodulatory effects due to the presence of other strains of bifidobacteria.
Intestinal barrier
Intestinal barrier dysfunction is an undervalued risk factor for CVD48. The integrity of the intestinal barrier is maintained by the connections between intestinal epithelial cells, which consist of proteins such as tight junction (TJ) proteins, adhesion junction (AJ) proteins, gap junction proteins, and desmosomal proteins49. It is essential for intracellular communication and paracellular permeability that TJ proteins are located at the top of the intestinal epithelial cells (Fig. 1c). Once the TJ proteins are impaired, bacteria and bacterial products such as lipopolysaccharide (LPS) in the intestines leak and enter the systemic circulation (Fig. 1a)48. There are accumulating reports on the association between the translocation of LPS and CVD. Abnormal elevation of serum LPS has been observed in AS, HP, chronic heart failure (CHF), and MI48. Indeed, impaired intestinal barrier, endothelial dysfunction, and pro-inflammatory response induced by LPS translocation have been shown to seriously affect the development of CVD.
It should be emphasized that the abundance of bifidobacteria directly impacts the integrity of the intestinal barrier. A previous study using mouse models demonstrated that a high-fat diet (HFD) damages the intestinal barrier by reducing the number of bifidobacterial50. The effects of bifidobacteria on TJ and AJ proteins have been demonstrated in recent studies. One study acknowledged that administration of bifidobacteria inhibited LPS translocation by increasing serum glucagon-like peptide-2 (GLP-2) to upregulate TJ proteins (occludin ZO1 and occludin) (Fig. 1a)17. As metabolites of bifidobacteria, short-chain fatty acids (SCFAs) are reported to protect intestinal barrier function by promoting assembly of TJ proteins and inhibiting activation of inflammatory vesicles (Fig. 1c). These data demonstrate that bifidobacteria is indeed beneficial in maintaining the intestinal barrier; and thus has the potential to be used in the repair of “leaky gut”. However, the precise mechanism underlying the protective impact of bifidobacteria on the intestinal barrier remains incompletely understood. This includes its interplay with structures like the villi and crypts of the intestinal epithelium, as well as whether changes in junction proteins are linked to the attenuation of inflammatory processes.
Gut microbiota metabolites
Intestinal microbial metabolites are novel risk factors for cardiovascular events51. A gastrointestinal tract is integral in digestion and absorption, and gut microbiota creates a bridge between diet and host. The gut microbiota produces an abundance of small-molecule metabolites when involved in the co-metabolism of food or exogenous substances, some of which are critical in the transmission of information between the host gut and distant organs, such as trimethylamine N-oxide (TMAO), SCFAs, conjugated fatty acids and secondary bile acids52,53. Recently, there has been a lot of interest on the causal relationship between the small molecules and CVD, and it has been found that these small-molecule metabolites play a role in the development and progression of CVD54.
Trimethylamine N-oxide (TMAO)
TMAO is a potential risk factor for chronic diseases, especially CVDs. The intake of red meat, milk, poultry, and eggs increases the body’s choline or trimethylamine products, which are metabolized by the intestinal microbiota to produce trimethylamine (TMA). TMA undergoes intestinal absorption and portal vein transport and is then oxidized to TMAO by flavin-containing monooxygenases (FMO) 1 and 3 in the liver (Fig. 1c)55. TMAO has been shown to be a potential promoter of CVD, especially AS56. Several mechanisms of TMAO-promoting AS have been proposed, which include promotion of the expression of inflammatory factors, breakdown of the balance of cholesterol metabolism, and induction of thrombosis57. It is worth mentioning that TMAO has been proven to predict the risk of CVD in multiple clinical cohorts, and has demonstrated uninterrupted prognostic value in a variety of adverse cardiac events (including coronary artery disease, HF, MI, death)58,59,60,61,62.
In addition, it has been hypothesized that bifidobacteria could down-regulate TMAO levels in vivo, which has been confirmed by findings from multiple studies. In the previous study on the mechanism of resveratrol against AS, it was demonstrated that the level of bifidobacteria in the intestinal microbiota of mice increased, accompanied by a decrease in the TMAO level63. This negative correlation between bifidobacteria and TMAO levels led to the exploration of more profound interactions. The TMAO-lowering properties of bifidobacteria have been explained by several mechanisms (Fig. 1c). A study conducted in choline mice showed that three bifidobacteria, including Bifidobacterium breve (B. breve) Bb4, B. longum BL1, and B. longum BL7, significantly decreased plasma TMAO levels and restored the abundance of some gut microbial species, while maintaining the activity of FMO18. This finding suggests that bifidobacteria may be reducing TMAO levels by modulating gut microbial abundance. Similarly, another study reported that administration of B. lactis F1-3-2 decreased plasma TMAO, which was not dependent on the regulation of FMO. Whether, it may have directly degraded TMA or adjusted the intestinal microbiota structure remains to be determined64. In addition, bifidobacteria may be antagonizing some strains responsible for synthesizing TMAO precursor molecules, such as Clostridium difficile65. Some probiotics have been proposed to reduce TMAO levels by altering miRNAs and modulating metabolomic profiles66, but data on bifidobacteria are still scarce and more targeted experimental evidence is needed.
Short-chain fatty acids (SCFAs)
Previously, data on the metabolism of SCFAs in CVD remained limited. However, with the growing evidence, it was shown that SCFAs are important in regulating cardiovascular functions67. In HP, both an increase in propionic and acetic acids induces vasodilation, and butyrate has been shown to relieve HP by inhibiting the angiotensin system in the kidney68. The properties of SCFAs such as immunomodulation, antioxidant stress, and improvement of lipid metabolism, are of great significance to the treatment of AS69,70. In addition, SCFAs act on the nervous system to protect the heart from injury and maintain its functions. A previous study showed that butyrate reversed autonomic imbalance in I/R rats and improved cardiovascular function by targeting the paraventricular nucleus and superior cervical ganglia71. It has also been established that butyrate may prevent ventricular arrhythmias after MI by inhibiting sympathetic remodeling72. SCFAs are a particular metabolite whose impact on the development of CVD is of far-reaching significance.
Human dietary fiber is degraded into organic acids by intestinal microflora, gas, and a large number of SCFAs73. Acetic acid, propionic acid, and butyric acid account for 90% of the SCFAs74. It is important to note that acetic acid is the main end-product of the metabolism of bifidobacteria. Bifidobacteria could indirectly increase butyric acid levels in the gut by cross-feeding interactions which enhance intestinal colonization of other butyric acid-producing commensal microorganisms (Fig. 1c), such as Faecalibacterium prausnitzii75 and Eubacterium hallii76. However, cross-feeding interactions between bifidobacteria and butyrate-producing bacteria vary from species to species, ranging from symbiotic to competitive relationships for energy substrates, which are largely related to the ability of bifidobacteria to degrade energy sources77. Currently, studies on the ability of different Bifidobacterium strains to degrade energy sources need to be deepened and continuously improved.
Indirect improvement of cardiovascular disease
Obesity
Obesity is associated with numerous severe health consequences, with cardiovascular disease (CVD) being a primary concern. It contributes to the development of CVD and increases CVD mortality78. Recent data suggest that severe obesity increases the risk of cardiovascular-related incidents in varying degrees: it increased the risk of HF by about 4-fold and increased the risk of coronary heart disease and stroke by nearly 2-fold79.
Appetite
Recent studies have suggested that bifidobacteria are involved in energy homeostasis and appetite regulation in the central nervous system (CNS) by improving levels of hormones such as leptin and gastrin80,81.
Leptin, a peptide hormone, is associated with the CNS’s perception of energy balance and food intake82. Leptin works by reducing dietary intake and body weight, so as the body fat increases, leptin continues to rise83. In chronically obese people, persistently had high levels of leptin in the blood reduce the sensitivity of leptin receptors in the hypothalamus, a phenomenon widely known as leptin resistance. Leptin resistance is characterized by a strong appetite, reduced energy expenditure and obesity. The mechanisms of leptin resistance include stimulation of inflammatory factors leading to abnormal signaling, reduced efficiency of blood-brain barrier transport, and receptor mutations. A study by Renata et al.81 demonstrated that after giving gastric gavage of probiotics containing B. bifidum to HFD-fed mice, the leptin resistance of obese mice was significantly improved, which was reflected in the significantly reduced food intake in mice. This may be due to the fact that probiotics containing B. bifidum significantly reduce mRNA levels of pro-inflammatory molecules TLR4 and IL-6 and expression of JNK and IKK proteins in the hypothalamus, thereby improving leptin signaling abnormalities (Fig. 2a). The molecular interaction between bifidobacteria and leptin remains unclarified. However, it has been demonstrated that SCFAs stimulate the expression of leptin in adipocytes by activating the free fatty acid receptor 3 (FFAR3)84, which provides a basis for future studies on the association between bifidobacteria and leptin.

a Bifidobacterium can control appetite by affecting the transmission of leptin and gastric hunger signaling pathways; it can also reduce obesity by inhibiting the disordered lipid metabolism process and reducing the impaired glucose metabolism in obese hosts in multiple ways. b Bifidobacterium can reduce the development of hyperlipidemia through cholesterol assimilation; high BSH ability to increase cholesterol excretion in the feces and limit cholesterol absorption, thus promote cholesterol conversion. c Bifidobacterium can reduce glucose metabolism by targeting hepatic gluconeogenesis genes, restoring insulin signaling pathway; producing SCFA to secrete GLP-1 and regulating islet beta cell growth and modulating the target inflammation to improve insulin resistance and reduce diabetes. (The figure does not contain any third-party material, the figure and each of the elements in the figure were created by the authors).
Besides reducing appetite by improving impaired leptin signaling pathways, bifidobacteria also improve ghrelin signaling pathways. Gastrin is an endocrine hormone produced in the gastric mucosa, which is responsible for the regulation of appetite and energy expenditure85,86. Specifically, ghrelin is secreted in large quantities during starvation, which then enters the blood circulation and reaches the center. Thereafter, it acts on the hypothalamic and midbrain limbic circuits to control dietary behavior and food intake87. For instance, a previous study showed that when non-obese individuals ate for 30 minutes, ghrelin was immediately suppressed by 39.5 % and continued to decline until it returned to baseline88. In contrast, for obese individuals, food intake did not seem to cause the ideal decrease in ghrelin88. Such postprandial ghrelin passivation is usually one of the reasons for excessive food intake in obese people. Previous studies using animal models showed that serum ghrelin levels were negatively correlated with the abundance of some bacterial populations in the gut microbiome, including the Bifidobacterium species89. Some scholars have explored the effect of bifidobacteria on human anti-obesity development. Findings from the studies showed that B. longum APC1472 significantly increased the ghrelin activity in obese individuals, which may demonstrate that B. longum APC1472 improves ghrelin signaling defects (Fig. 2a)80.
Lipid and glucose metabolism
Apart from the appetite in obese people, metabolic abnormalities directly enhance the process of obesity. It has been shown that obesity-related metabolic disorders are often accompanied by a decrease in the genus Bifidobacterium in the intestines90. Nevertheless, accumulating experimental evidence suggests that bifidobacteria supplementation is beneficial in inhibiting lipid metabolism process in obese host disorders in multiple ways.
In general, in non-obese people, there is maintenance of a dynamic balance between fat consumption and production. However, due to long-term unhealthy lifestyles or some disease factors, there is destruction of the lipid homeostasis in the body, and the decomposition of fat is far less than the production of fat, resulting in excessive accumulation of lipids and eventual obesity91. A variety of bifidobacteria, such as Bifidobacterium adolescentis (B. adolescentis) and Bifidobacterium animalis subsp. animalis (B. animalis), showed a tendency to restore lipid profiles (including LDL-C, HDL-C, TC, triglyceride (TG)) to normal levels19,92. It has been demonstrated that bifidobacteria regulate fat accumulation by upregulating mRNA expression of thermogenic genes and lipolytic enzymes, and inhibiting activation of lipogenic genes. For example, B. adolescentis was observed to upregulate mRNA expression of Ucp-1, Pgc1-α, Ppar-γ, and Ppar-α genes in HDF-fed mice, to produce fatty acids which promote lipid metabolism in brown adipose tissue (Fig. 2a)19. Ppar-α upregulates the expression of enzymes responsible for mitochondrial fatty acid oxidation, while Ppar-γ activation triggers brown adipocyte differentiation and adipogenesis93. These results suggest that acceleration of fatty acid conversion is one of the ways bifidobacteria use to improve fat metabolism (Fig. 2). In addition, some bifidobacteria may also improve lipid metabolism by producing EPS, a substance with potential health benefits94. Another study demonstrated that the expression of acyl-CoA oxidase 1 (ACOX 1), carnitine palmitoyl transferase 1A (CPT 1A), and 3-hydroxy-3-methyl-3-glutaryl-CoA reductase (Hmgcr) of Diet-Induced Obese Mice was significantly upregulated by the application of B. animalis IPLA R1, which produce EPS (Fig. 2a)92. ACOX 1 and CPT 1A are rate-limiting enzymes in the fatty acid oxidation pathway, while Hmgcr is a rate-limiting enzyme in cholesterol synthesis95. These data suggest that B. animalis IPLA R1 can inhibit fat accumulation in the liver by accelerating lipid oxidation and increasing cholesterol excretion (Fig. 2a). Similarly, SCFAs, as intestinal microbial metabolites, can be directly involved in improving lipid metabolism because of their ability to increase lipid oxidation (Fig. 2a)69,96.
Numerous studies have indicated that the regulation of lipid metabolism by bifidobacteria is often accompanied by the normalization of blood glucose levels97,98,99. Parallel to lipid metabolism disorders, glucose metabolism disorders are also closely related to obesity. A study by Cano PG et al.99 found that seven weeks of continuous administration of Bifidobacterium pseudocatenulatum (B. pseudocatenulatum) CECT 7765 reduced blood glucose levels in obese HFD-fed mice by 17%, demonstrating its ability to increase glucose tolerance. The improvement of glucose metabolism parameters by bifidobacteria may be associated with the activation of glucokinase (GCK) (Fig. 2a)100. It is worth noting that GCK has better control over the rate of insulin secretion in pancreatic β-cells, and thus small changes in its activity can cause fluctuations in the insulin secretion threshold and affect glucose homeostasis100. Previous systematic analyses showed that yogurt fermented B. longum 070103 successfully activates GCK and thus significantly reduces fasting blood glucose and improves glucose tolerance as well as insulin resistance101. Moreover, specific regulation of the abundance of crucial intestinal microbes, reduction of the levels of metabolites such as 3-indolyl sulfate and 4-hydroxybutyric acid to alleviate glucose metabolic disorders could be included in the mechanism of bifidobacteria hypoglycemic activities101.
Hyperlipidaemia (HLP)
HLP is defined as excessive blood lipids (the sum of all lipids in the plasma) due to abnormal fat metabolism or transport. Abnormal levels of one or more lipids in the plasma often lead to HLP, and include increased total cholesterol (TC), TG, low-density lipoprotein cholesterol (LDL-C), and decreased high-density lipoprotein cholesterol (HDL-C)102. This explains why HLP presents as hypercholesterolemia, hypertriglyceridemia, or both (mixed HLP). However, lack of intervention eventually leads to CVD, regardless of the kind of HLP. Early intervention of dyslipidemia is essential for primary and secondary prevention of CVD, especially AS and coronary heart disease.
Hypercholester
Cholesterol is one of the subtypes of lipids which is transported to the whole body in form of lipoproteins. Lipoproteins can be divided into LDL-C, very low-density lipoprotein cholesterol (VLDL-C), and HDL-C, which perform unique functions in the body. LDL-C carries cholesterol into the artery and binds to macrophages on the arterial wall. Macrophages absorb cholesterol and gradually develop into foam cells, burying the hidden danger of AS. LDL-C is, therefore, generally considered “bad” cholesterol. On the contrary, HDL-C transports cholesterol accumulated in the arteries to the liver for metabolism, thereby preventing CVD. It has been suggested that every 10 mmol/L reduction in LDL cholesterol reduces the risk of CVD and mortality by 22%102. Adverse reactions or contraindications to statins and fibrates currently used in clinical practice are plaguing doctors and patients103. The role of bifidobacteria in lowering cholesterol is constantly emphasized, not only because of its effectiveness but also its diverse mechanisms of action in lowering the cholesterol. Cholesterol assimilation occurs when Bifidobacterium binds cholesterol to the surface of the cell and absorbs it into the membrane’s phospholipid bilayer (Fig. 2b)104. Different Bifidobacterium strains showed varying degree of assimilation abilities. For example, under the same experimental conditions, the adsorption rate of Bifidobacterium bifidum (B. bifidum) MB109 to cholesterol was 52%, while that of Bifidobacterium longum subsp. infantis (B. infantis) ATCC 15697 was 34%105. Secondly, bifidobacteria’s bile salt hydrolase (BSH) activity enables it to reduce cholesterol by participating in the metabolism of the bile acids. More specifically, Bifidobacterium hydrolyzes conjugated bile acids into primary bile acids through its high BSH capacity, making them more easily excreted in feces (Fig. 2b). The reduction of bile salts and the loss of bile acids stimulate the liver to increase cholesterol conversion, which decreases serum cholesterol levels. These findings were confirmed by Al-Sheraji and Jiang in an in vivo study using rats20,106. BSH activity has also been shown to limit cholesterol absorption by reducing the solubility of bile acids (Fig. 2b)107. Thirdly, cholesterol conversion also contributes to the removal of cholesterol by bifidobacteria (Fig. 2b). Another study observed an increase in coprostanol in cholesterol-rich cells treated with B. bifidum PRL2010, accompanied by upregulation of BBPR 0519, which is predicted to encode the aldehyde/ketone reductase which catalyzes the conversion of cholesterol to coprostanol108.
High triglycerides (TG)
More than 30 years ago, the negative effect of high TG to CVD was comparable with that of high TC109. Although the advent of statins has led to more focus on lowering LDL-C over time, there has been a renewed interest in TG110. It has been proved that the size of a lipoprotein can enter the arterial wall when TG concentration is slightly or moderately elevated (2–10 mmol/L), thus increasing the risk of CVD111,112. In contrast, when its concentration is seriously elevated (>50 mmol/L), the lipoproteins become too large to enter the arterial wall, which affect people’s judgment of the TG113. Although there may be more than one mechanism for TG in CVD, there is one mechanism that has been discussed most clearly: high TG leads to increased concentrations of residual cholesterol, which enters the arteries, promotes inflammatory episodes and foam cell formation, which ultimately increases the risk of CVD and death114.
Bifidobacteria were negatively correlated with TG levels. Further clinical studies have shown that supplementation with bifidobacteria or intake of functional foods containing bifidobacteria could reduce serum TG levels115,116. An improvement in the overall lipid profile usually accompanies TG-lowering effect of bifidobacteria. The TG-lowering mechanism of the bifidobacteria is less precise than that of cholesterol-lowering. However, the benefits of bifidobacteria and good tolerance in reducing TG make it worthy of consideration in the therapy of lipid abnormalities.
Type 2 diabetes mellitus (T2DM)
Diabetes is an established risk factor for CVD such as HF, coronary heart disease, stroke, and atrial fibrillation117. It is estimated that 2–4 times more diabetic patients die from CVD than non-diabetic patients, and at least 68% of diabetics over 65 years old die from various forms of CVD118. More than two-thirds of patients with T2DM are reported to have HP119. According to a meta-analysis report, the risk of HF was 1.11 (95% CI, 1.04–1.17) for every 1 mmol/L (≈18 mg/dL) increase in fasting blood glucose, indicating a positive correlation between fasting blood glucose and HF120.
Recently, the unique benefits of bifidobacteria in the treatment of T2DM have been validated in both animal studies and clinical trials. More studies have revealed the mechanisms underlying this beneficial role, and several phased results have been achieved. Studies have claimed that bifidobacteria have a significant effect on improving insulin resistance (IR) in patients with T2DM, and some strains are even more effective than metformin21,29,121,122. The experimental studies by Zhang et al.29 showed that bifidobacteria utilize several pathways to improve insulin sensitivity. First, bifidobacteria may alleviate IR by targeting hepatic gluconeogenesis genes to reduce gluconeogenesis (Fig. 2c). The study used B. animalis 01 in T2DM rats, and showed that B. animalis 01 down-regulated phosphoenolpyruvate carboxykinase (PEPCK), glucose-6-phosphatase (G6Pase), and upregulated the expression of Nrf2, IRS-2, PI3K, and AKT-2 genes (Fig. 2c)29. Researchers have suggested that B. longum BL12 and B. lactis HY8101 down-regulate PEPCK and G6Pase in the liver of T2DM mice21,121. Nrf2 is a protective factor against oxidative damages. Activation of the Nrf2 inhibits IRS-2 phosphorylation and thus increases the expression of downstream signals such as PI3K and AKT (Fig. 2c). The IRS – PI3K – AKT pathway is critical in hepatic insulin signaling. Therefore, enhancement of the antioxidant capacity is another effective way adopted by bifidobacteria to restore insulin signaling and repair IR.
Inflammation is known to be associated with induction of IR and development of T2DM123. Due to its anti-inflammatory properties, B. adolescentis strains have recently been found to alleviate IR. The diabetic state of T2DM mice was improved after administration of B. adolescentis. The pro-inflammatory factors including TNF-α, IL-6, and IFN-γ were significantly inhibited, and the concentrations of butyric and propionic acids were significantly increased (Fig. 2c)122. Based on the high association between the elevated concentrations of SCFAs caused by these strains and their effects on blood glucose concentrations, Qian122 hypothesized that the ameliorative diabetic effects of adolescent bifidobacteria are mediated through the bifidobacteria-gut microbiota-SCFAs-inflammation axis. B. lactis GCL2505, which has been evaluated for improving diabetes, appears to regulate SCFAs (particularly acetate) levels124. In addition, elevated SCFAs stimulate GPR43, leading to the secretion of GLP-1, which regulates β-cell growth, stimulates glucose-dependent insulin release, and inhibits glucagon secretion (Fig. 2c)124.
Current applications in cardiovascular disease
Clinical studies on applications of Bifidobacterium species in CVD are summarized in Table 1. The studies vary by subject, sample size, bacteria, product, dosage, and study design.
Hypertension (HP)
HP is a chronic disease that is characterized by continuously high arterial blood pressure levels. Long-term HP is the most significant risk factor for coronary artery disease, stroke, HF, atrial fibrillation, and other CVDs. A 20 mmHg increase in systolic blood pressure and a 10 mmHg increase in diastolic blood pressure are associated with a two-fold increase in the risk of death from stroke, heart disease, or other vascular diseases125. Based on evidence from CVD attribution analyses, a rightward shift in blood pressure distribution resulting in major cardiovascular diseases in humans has been proposed126.
A link between reduced bifidobacteria abundance and increased blood pressure incidences in children with type 1 diabetes mellitus (T1DM) has been proposed13. In this study, children were assigned into three groups: healthy control group (HC, n = 5), T1DM group with normal blood pressure levels (T1DM-Normo, n = 17), and type 1 diabetes group with elevated blood pressure levels (T1DM-HBP, n = 7)13. Analysis of gut microbiota for each group revealed that bifidobacteria abundance in the guts of children in the T1DM-HBP group were significantly lower than in the other two groups13. Another trial indicated that supplementation with probiotics containing B. lactis HN019 reduced systolic blood pressure levels by 5 mmHg and diastolic blood pressure levels by 2 mmHg in women with arterial HP, with effective improvements in lipid metabolism as well as fasting glucose levels127.
In Germany, 100 grade 1 HP patients were invited to participate in probiotic intervention trials128. After 8 weeks of dynamic nocturnal blood pressure monitoring and evaluation of fecal microbiome composition as well as immune cell phenotypes, it was proposed that the mechanisms by which bifidobacteria reduce HP involve improving immune cell homeostasis by transforming dietary metabolic components128. This hypothesis has been tested in rat models. In deoxycorticosterone acetate (DOCA) salt hypertensive rat models, B. breve CECT7263 alleviated HP by increasing acetate concentrations in the gut and reducing TMA, restoring Th17/Treg immune homeostasis and suppressing vascular NADPH oxidase activities129. In spontaneously hypertensive rat (SHR) models, B. breve CECT7263 suppressed the elevated blood pressure levels by increasing the number of butyrate-producing bacteria, preventing Th17/Treg dysregulation, reducing endotoxemia, and improving endothelial dysfunctions130.
Atherosclerosis (AS)
Clinically, AS is an intimal disease in which fatty deposits form plaques in inner layers of arteries131. Then, growth of plaques leads to thrombosis and bulging in arteries. Fibrous tissue proliferation and calcium deposition accelerates arterial wall thickening and hardening131. Finally, the narrowing or blockage of arterial lumen leads to ischemia and necrosis of tissues and organs supplied by the artery131. Physiologically, the processes involved in AS development are complex and slow, involving many pathological changes, including hypercholesterolemia, inflammation, oxidative stress, and TMAO66,131,132. Even though the cholesterol-lowering, anti-inflammatory, anti-oxidative stress, and TMAO-modulating effects of bifidobacteria have been previously reported, the corresponding mechanisms are described in detail in Milad’s review133. However, more preclinical and clinical trials should be performed to improve AS through bifidobacteria supplementation.
Randomized trials involving individuals with mild to moderate hypercholesterolemia revealed significant cholesterol-lowering effects of bifidobacterial134. Thirty-two adult males were randomized into two groups and administered with 3 × 100 ml/day of ordinary yogurt or fermented yogurt for 4 weeks116. A decrease in TC levels was established in the B. longum BL1 fermented yogurt group and the effects were more pronounced in individuals with moderate hypercholesterolemia (TC > 240 mg/dl)116. Another randomized trial revealed that in mild hypercholesterolemia patients with TC levels between 180 and 220 mg/dl, 10 weeks of a formula containing B. lactis Bb12 reduced their TC and LDL-C levels by 8.1% and 10.4%, respectively135. A clinical trial involving healthy young people showed that bifidobacteria improved lipoprotein profiles in hypercholesterolemia patients but not normal cholesterol levels136. In addition, some valuable clinical data showed that bifidobacteria improves TC levels T2DM patients137,138.
An animal study investigating the effects of probiotics on plasma TMAO revealed that 7 of 16 Bifidobacterium strains significantly reduced plasma TMAO concentrations, and that the plasma TMAO reduction rate for B. longum BL1 was as high as 30.89%18. Current clinical trials have shown that bifidobacteria inhibits TMAO levels. In a previous study, TMAO levels were found to be elevated in 40 healthy young men (20–25 years) that had been subjected to a phosphatidylcholine challenge test139. Then, they were randomized into two groups, one of which received probiotic intervention139. The TMAO levels for most participants in the probiotic group decreased, however, the decrease was insignificant139. A similar randomized double-blind trial involving 27 healthy volunteers (mean age 47.1 years) obtained contrasting results140. In this study, participants were assigned to receive either B. lactis LKM512 or placebo140. After 12 weeks, fecal TMA concentrations and the abundance of TMA-producing bacteria were significantly low in the bifidobacteria group, relative to the placebo group (p < 0.05)140. It is important to note that since there is a positive correlation between plasma TMAO levels and age, age of the subjects may have been a critical factor leading to different outcomes of the two experiments141,142. This may be due to the fact that expressions of FMO3 isoforms increased with age in clinical models, which promoted TMA transformation to TMAO143. Overall, studies should elucidate on the link between bifidobacteria and TMAO.
The potential mechanisms by which bifidobacteria affect atherosclerotic CVD via low-grade inflammation have been discussed144. In the past 5 years, clinical trials have reported the positive effects of bifidobacteria on inflammation and oxidative stress in diabetic patients with coronary heart disease145,146. One of the trials involved 60 overweight, diabetic, and coronary heart disease patients aged 50–58 years and it aimed at assessing the effects of synbiotics (including bifidobacteria) on inflammatory biomarkers of carotid intima-media thickness and oxidative stress146. After 12 weeks of treatment, bifidobacteria significantly reduced hsCRP as well as plasma malondialdehyde levels, and significantly increased nitric oxide (NO) levels146. In Poland, probiotic supplements containing various Bifidobacterium strains were shown to reduce IL-6, TNF-α, and thrombomodulin levels in postmenopausal obese women and effectively improved arterial stiffness147.
Myocardial infarction (MI)
Clinically, MI, known as myocardial ischemic necrosis, involves a rapid reduction or interruption of coronary blood supply due to coronary arterial disease (such as AS, spasm), leading to acute and persistent myocardial ischemia in the myocardium of coronary supply sites, and ultimately resulting in myocardial necrosis.
Previously, studies have reported a low gut abundance of bifidobacteria in rats148 and humans149 with MI. However, they did not conclusively determine whether bifidobacteria are cardioprotective against MI. The direct association between bifidobacteria and MI was first reported by Lam et al. in 2012. Lam et al.150 reported that myocardial infarct sizes were reduced by 29% in MI rats that had been fed on Goodbelly for 14 days. Goodbelly is a commercially available probiotic juice containing two probiotics, Lactobacillus plantarum 299v and B. lactis Bi-07. Due to the absence of relevant evidence, it has been postulated that protective effects of this probiotic are as a result of reduction in serum leptin levels. Danilo et al.151 found that administration of B. lactis B420 for 4 weeks or 7 days significantly mitigated myocardial infarct sizes in mice following I/R. The epigenetic mechanisms of B. lactis B420 against MI were also identified. Further, Danilo et al. explained that the anti-MI effects of B. lactis B420 were achieved by suppressing the levels of inflammatory factors and accelerating transitions to M2-type macrophages via the mediatory effects of anti-inflammatory T-regulatory immune cells. Their findings are in tandem with those of Jafar Sadeghzadeh et al. who found that oral administration of a probiotic combination formulation containing B. breve exerted cardioprotective effects on rats with infarct-like myocardial injuries by attenuating TNF-α and inhibiting oxidative stress152.
Depression-like behaviors and development of depression are frequent in MI patients153. Bifidobacteria have been shown to reduce the development of depression-like behaviors after MI in animal models154,155,156. Since increased apoptosis was observed in various limbic system structures after MI to varying degrees, Girard et al.157 first proposed that prophylactic supplementation with B. longum and Lactobacillus reduces apoptosis. They confirmed this hypothesis in rat models and postulated that this combination of probiotics exerts anti-inflammatory effects leading to inhibition of apoptosis in the limbic system157. In addition to preventive effects, probiotic supplementation after ischemia-reperfusion has been shown to maintain its beneficial effects while improving depression-like behaviors after MI155. Later, scholars determined that B. longum plays more beneficial roles in probiotic combinations to improve depression after MI156. A randomized, double-blind, placebo-controlled clinical trial revealed the beneficial effects of probiotic supplementation (Lactobacillus rhamnosus) on depressive symptoms, inflammation, and oxidative stress in MI patients158. Bifidobacteria, which has shown excellent anti-depressive effects after MI in preclinical studies, may also hold up in clinical trials, although the link has yet to be established.
Heart failure (HF)
Since HF is not an independent disease, its definition in academia is inconclusive. The American Heart Association defines it as a complex clinical syndrome resulting from various cardiac structural or functional diseases that impair ventricular filling or ejection capacities159. Long-term HP increases cardiac load, leading to myocardial hypertrophy, remodeling, and HF160. Coronary atherosclerotic heart disease (coronary heart disease) results in long-term heart ischemia and hypoxia, leading to gradual weakening of heart contractions, thereby inducing HF161. Physiologically, MI is a risk factor for HF in coronary heart disease. This is because MI leads to a sharp decrease in myocardial contractility and to a significant decrease in cardiac pumping volume. Thus, after infarction, the myocardium becomes fragile, aggravating HF development162. HF is usually the common end for multiple CVDs163. The 5-year survival rate for HF patients is only 45%, indicating that new prevention and treatment strategies are urgently required to improve on its prognosis164.
Soluble tumor necrosis factor-like weak inducer of apoptosis (sTWEAK) is an independent predictor of mortality risk in CHF, and non-ischemic HF, implying that it is a potential new cardiovascular biomarker165,166,167. Compared with healthy individuals, the concentrations of sTWEAK in patients with congestive HF, coronary artery disease, and AS have been shown to be suppressed167. A triple-blind clinical trial randomly assigned 90 CHF patients into two groups receiving probiotic yogurt containing bifidobacteria or ordinary yogurt for 10 weeks168. Compared with ordinary yogurt, probiotic yogurt increased serum sTWEAK levels in CHF patients168. The increase in serum sTWEAK levels may have been because probiotic intake reduces inflammation, thereby downregulating Fn14, the only receptor for sTWEAK, which prevents sTWEAK from binding Fn14. In addition, a randomized triple-blind clinical trial showed that probiotic yogurt significantly reduced serum ox-LDL levels, which might be related to the increase in total antioxidant capacities through multiple pathways (production of antioxidant metabolites, modification of MAPK, NF-κB, and other pathways, as well as regulation of ROS-producing enzymes)169.
In addition to patients with HF alone, a previous clinical trial focused on gastric cancer patients with coronary heart disease and HF complications because apparent intestinal disorders are consistently observed in such populations and they stimulate HF progression170. In patients with gastric cancer with coronary heart disease and HF, probiotic capsules containing B. longum enhanced the intestinal barrier and corrected intestinal microbial imbalance as well as HF, thereby promoting patient rehabilitation170.
Several clinical trials have reported the positive effects of bifidobacteria in HF patients. However, these clinical trials used probiotic combinations containing bifidobacteria rather than individual Bifidobacterium strains alone, which blurred the role of bifidobacteria to some extent. Preclinical and clinical studies should be performed to elucidate on the independent beneficial effects of bifidobacteria in HF patients.
Prospects
This review has elucidated the critical role of bifidobacteria in the prevention of CVD development. On the one hand, bifidobacteria prevented damage to cardiac functions through the maintenance of intestinal barrier function and its antioxidant and immunomodulatory effects directly. On the other hand, bifidobacteria can also indirectly reduce the cardiac burden of obesity, HLP, and T2DM by affecting hormone secretion, regulating metabolism, assimilating cholesterol, and improving IR. In addition, bifidobacteria have found extensive application and demonstrated promising progress in various classic CVDs, such as HP, AS, MI, and HF. Indeed, some unique metabolic functions of Bifidobacterium may play a non-negligible role in cardiovascular disease, such as the metabolism of tryptophan, the production of its indole derivatives, and the regulation of peripheral serotonin, but the lack of evidence leads to the need for more attention and in-depth exploration.
It is important to recognize that advances in the field of molecular biology and genetic engineering techniques will pave the way for in-depth studies into all probiotics, including bifidobacteria. In recent years, several mutagenesis methods have been introduced in the field of bifidobacteria, including homologous recombination systems with non-replicating plasmids or temperature-sensitive plasmids, random rotor-based mutagenesis systems, and inducible plasmid self-destruction-assisted systems171,172,173,174. Although these strategies have some shortcomings, there are efforts to circumvent these problems through strategies, such as the development of endogenous CRISPR-Cas systems171. On the one hand, recent advances in the field of molecular biology and genetic engineering techniques have facilitated a deeper understanding of bifidobacteria-host interactions175. For example, by analyzing the genome sequence and in vivo transcriptome studies of Bifidobacterium breve, O’Connell176 identified a key site that influences its colonization of the mouse host intestine and highlighted that the ability of different strains to colonize the intestine is highly correlated with their ability to utilize carbohydrates. Furthermore, genetic engineering holds profound significance for forthcoming therapeutic advancements and industrial applications involving bifidobacteria. By harnessing sophisticated genetic engineering methods, the potential exists to enhance stress resilience, engineer targeted delivery systems to combat pathogens, eradicate antibiotic resistance, achieve luciferase labeling, and even generate entirely novel transgenic bifidobacterial strains that surpass wild-type counterparts in both efficacy and safety177, which can help develop novel therapeutic approaches and facilitate further clinical optimization.
In summary, addressing and controlling the bifidobacterial composition holds promise for advancing innovative therapeutic strategies in the context of CVD. The manipulation of the Bifidobacterium genome via genetic engineering to create pertinent foods and medications is anticipated to pave the way for safer and more efficient utilization of Bifidobacterium in the future, augmenting its potential as a treatment for CVD in human populations.
References
-
Mendis, S., Puska, P., Norrving, B. & Organization, W. H. Global Atlas on Cardiovascular Disease Prevention and Control (World Health Organization, 2011).
-
Organization, W. H. WHO Cardiovascular Diseases (CVDs) 2021, http://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (2021).
-
Jin, J. Counseling on healthy living to prevent cardiovascular disease in adults without risk factors. Jama 318, 210 (2017).
Google Scholar
-
Sack, M. N., Fyhrquist, F. Y., Saijonmaa, O. J., Fuster, V. & Kovacic, J. C. Basic biology of oxidative stress and the cardiovascular system: part 1 of a 3-part series. J. Am. Coll. Cardiol. 70, 196–211 (2017).
Google Scholar
-
Papadopoulos, P. D. et al. The emerging role of the gut microbiome in cardiovascular disease: current knowledge and perspectives. Biomedicines 10, https://doi.org/10.3390/biomedicines10050948 (2022).
-
Jin, L. et al. Gut microbes in cardiovascular diseases and their potential therapeutic applications. Protein Cell 12, 346–359 (2021).
Google Scholar
-
Alessandri, G., van Sinderen, D. & Ventura, M. The genus Bifidobacterium: from genomics to functionality of an important component of the mammalian gut microbiota running title: Bifidobacterial adaptation to and interaction with the host. Comput. Struct. Biotechnol. J. 19, 1472–1487 (2021).
Google Scholar
-
Li, J. et al. Bifidobacterium: a probiotic for the prevention and treatment of depression. Front. Microbiol. 14, 1174800 (2023).
Google Scholar
-
Bottacini, F., Ventura, M., van Sinderen, D. & O’Connell Motherway, M. Diversity, ecology and intestinal function of bifidobacteria. Micro. Cell Fact. 13, S4 (2014).
Google Scholar
-
Harmsen, H. J. et al. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol. Nutr. 30, 61–67 (2000).
Google Scholar
-
Odamaki, T. et al. Age-related changes in gut microbiota composition from newborn to centenarian: a cross-sectional study. BMC Microbiol. 16, 90 (2016).
Google Scholar
-
Turroni, F., Ribbera, A., Foroni, E., van Sinderen, D. & Ventura, M. Human gut microbiota and bifidobacteria: from composition to functionality. Antonie Van Leeuwenhoek 94, 35–50 (2008).
Google Scholar
-
Lakshmanan, A. P. et al. Bifidobacterium reduction is associated with high blood pressure in children with type 1 diabetes mellitus. Biomed. Pharmacother. 140, 111736 (2021).
Google Scholar
-
Liang, X. et al. Krill oil combined with Bifidobacterium animalis subsp. lactis F1-7 alleviates the atherosclerosis of ApoE(-/-) mice. Foods 10, https://doi.org/10.3390/foods10102374 (2021).
-
Bernini, L. J. et al. Beneficial effects of Bifidobacterium lactis on lipid profile and cytokines in patients with metabolic syndrome: a randomized trial. Effects of probiotics on metabolic syndrome. Nutrition 32, 716–719 (2016).
Google Scholar
-
Wang, Y. et al. Supernatants of Bifidobacterium longum and Lactobacillus plantarum strains exhibited antioxidative effects on A7R5 cells. Microorganisms 9, https://doi.org/10.3390/microorganisms9020452 (2021).
-
Cani, P. D. et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 58, 1091–1103 (2009).
Google Scholar
-
Wang, Q. et al. Bifidobacterium breve and Bifidobacterium longum attenuate choline-induced plasma trimethylamine N-oxide production by modulating gut microbiota in mice. Nutrients 14, https://doi.org/10.3390/nu14061222 (2022).
-
Wang, B. et al. Bifidobacterium adolescentis isolated from different hosts modifies the intestinal microbiota and displays differential metabolic and immunomodulatory properties in mice fed a high-fat diet. Nutrients 13, https://doi.org/10.3390/nu13031017 (2021).
-
Jiang, J. et al. Strain-specific effects of Bifidobacterium longum on hypercholesterolemic rats and potential mechanisms. Int. J. Mol. Sci. 22, https://doi.org/10.3390/ijms22031305 (2021).
-
Hao, J. et al. The antidiabetic effects of Bifidobacterium longum subsp. longum BL21 through regulating gut microbiota structure in type 2 diabetic mice. Food Funct. 13, 9947–9958 (2022).
Google Scholar
-
Lorenzon Dos Santos, J., Quadros, A. S., Weschenfelder, C., Garofallo, S. B. & Marcadenti, A. Oxidative stress biomarkers, nut-related antioxidants, and cardiovascular disease. Nutrients 12, https://doi.org/10.3390/nu12030682 (2020).
-
Averina, O. V., Poluektova, E. U., Marsova, M. V. & Danilenko, V. N. Biomarkers and utility of the antioxidant potential of probiotic Lactobacilli and Bifidobacteria as representatives of the human gut microbiota. Biomedicines 9, https://doi.org/10.3390/biomedicines9101340 (2021).
-
Wang, W. & Kang, P. M. Oxidative stress and antioxidant treatments in cardiovascular diseases. Antioxidants 9, https://doi.org/10.3390/antiox9121292 (2020).
-
Hausenloy, D. J. & Yellon, D. M. Myocardial ischemia-reperfusion injury: a neglected therapeutic target. J. Clin. Invest. 123, 92–100 (2013).
Google Scholar
-
Lu, W. et al. Bifidobacterium longum CCFM752 prevented hypertension and aortic lesion, improved antioxidative ability, and regulated the gut microbiome in spontaneously hypertensive rats. Food Funct. 13, 6373–6386 (2022).
Google Scholar
-
Dev, K. et al. Dietary Mannan-oligosaccharides potentiate the beneficial effects of Bifidobacterium bifidum in broiler chicken. Lett. Appl. Microbiol. 71, 520–530 (2020).
Google Scholar
-
Xu, S., Chen, W., Xie, Q. & Xu, Y. Obacunone activates the Nrf2-dependent antioxidant responses. Protein Cell 7, 684–688 (2016).
Google Scholar
-
Zhang, J. et al. Anti-diabetic effects of Bifidobacterium animalis 01 through improving hepatic insulin sensitivity in type 2 diabetic rat model. J. Funct. Foods 67, 103843 (2020).
Google Scholar
-
Aragozzini, F., Ferrari, A., Pacini, N. & Gualandris, R. Indole-3-lactic acid as a tryptophan metabolite produced by Bifidobacterium spp. Appl. Environ. Microbiol. 38, 544–546 (1979).
Google Scholar
-
Ehrlich, A. M. et al. Indole-3-lactic acid associated with Bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 20, 357 (2020).
Google Scholar
-
Li, S. et al. Bifidobacterium longum R0175 protects mice against APAP-induced liver injury by modulating the Nrf2 pathway. Free Radic. Biol. Med. 203, 11–23 (2023).
Google Scholar
-
Wang, Y. et al. Antioxidant properties of probiotic bacteria. Nutrients 9, https://doi.org/10.3390/nu9050521 (2017).
-
Mahdavi-Roshan, M., Salari, A., Kheirkhah, J. & Ghorbani, Z. The effects of probiotics on inflammation, endothelial dysfunction, and atherosclerosis progression: a mechanistic overview. Heart Lung Circ. 31, e45–e71 (2022).
Google Scholar
-
Fonseca, F. A. & Izar, M. C. Role of inflammation in cardiac remodeling after acute myocardial infarction. Front Physiol. 13, 927163 (2022).
Google Scholar
-
He, S. et al. Gut intraepithelial T cells calibrate metabolism and accelerate cardiovascular disease. Nature 566, 115–119 (2019).
Google Scholar
-
Alessandri, G., Ossiprandi, M. C., MacSharry, J., van Sinderen, D. & Ventura, M. Bifidobacterial dialogue with its human host and consequent modulation of the immune system. Front. Immunol. 10, 2348 (2019).
Google Scholar
-
Ruiz, L., Delgado, S., Ruas-Madiedo, P., Sánchez, B. & Margolles, A. Bifidobacteria and their molecular communication with the immune system. Front. Microbiol. 8, 2345 (2017).
Google Scholar
-
Zelante, T., Fric, J., Wong, A. Y. & Ricciardi-Castagnoli, P. Interleukin-2 production by dendritic cells and its immuno-regulatory functions. Front. Immunol. 3, 161 (2012).
Google Scholar
-
Flint, H. J., Duncan, S. H., Scott, K. P. & Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 74, 13–22 (2015).
Google Scholar
-
Fang, Z. et al. Bifidobacterium longum mediated tryptophan metabolism to improve atopic dermatitis via the gut-skin axis. Gut Microbes 14, 2044723 (2022).
Google Scholar
-
Qi, Q. et al. Host and gut microbial tryptophan metabolism and type 2 diabetes: an integrative analysis of host genetics, diet, gut microbiome and circulating metabolites in cohort studies. Gut 71, 1095–1105 (2022).
Google Scholar
-
Imamdin, A. & van der Vorst, E. P. C. Exploring the role of serotonin as an immune modulatory component in cardiovascular diseases. Int. J. Mol. Sci. 24, https://doi.org/10.3390/ijms24021549 (2023).
-
Engevik, M. A. et al. Human-derived Bifidobacterium dentium modulates the mammalian serotonergic system and gut-brain axis. Cell Mol. Gastroenterol. Hepatol. 11, 221–248 (2021).
Google Scholar
-
Wang, Y. et al. Probiotics and fructo-oligosaccharide intervention modulate the microbiota-gut brain axis to improve autism spectrum reducing also the hyper-serotonergic state and the dopamine metabolism disorder. Pharm. Res. 157, 104784 (2020).
Google Scholar
-
Tatsuoka, M. et al. Consumption of indigestible saccharides and administration of Bifidobacterium pseudolongum reduce mucosal serotonin in murine colonic mucosa. Br. J. Nutr. 127, 513–525 (2022).
Google Scholar
-
Taverniti, V. et al. Probiotics modulate mouse gut microbiota and influence intestinal immune and serotonergic gene expression in a site-specific fashion. Front. Microbiol. 12, 706135 (2021).
Google Scholar
-
Lewis, C. V. & Taylor, W. R. Intestinal barrier dysfunction as a therapeutic target for cardiovascular disease. Am. J. Physiol. Heart Circ. Physiol. 319, H1227–h1233 (2020).
Google Scholar
-
Violi, F. et al. Gut-derived low-grade endotoxaemia, atherothrombosis and cardiovascular disease. Nat. Rev. Cardiol. https://doi.org/10.1038/s41569-022-00737-2 (2022).
-
Cani, P. D. et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57, 1470–1481 (2008).
Google Scholar
-
Heianza, Y., Ma, W., Manson, J. E., Rexrode, K. M. & Qi, L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: a systematic review and meta-analysis of prospective studies. J. Am. Heart Assoc. 6, https://doi.org/10.1161/jaha.116.004947 (2017).
-
Tang, W. H., Kitai, T. & Hazen, S. L. Gut microbiota in cardiovascular health and disease. Circ. Res. 120, 1183–1196 (2017).
Google Scholar
-
Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 449, 819–826 (2007).
Google Scholar
-
Tang, W. H. W., Li, D. Y. & Hazen, S. L. Dietary metabolism, the gut microbiome, and heart failure. Nat. Rev. Cardiol. 16, 137–154 (2019).
Google Scholar
-
Hoyles, L. et al. Metabolic retroconversion of trimethylamine N-oxide and the gut microbiota. Microbiome 6, 73 (2018).
Google Scholar
-
Brown, J. M. & Hazen, S. L. The gut microbial endocrine organ: bacterially derived signals driving cardiometabolic diseases. Annu Rev. Med. 66, 343–359 (2015).
Google Scholar
-
Zhu, W. et al. Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell 165, 111–124 (2016).
Google Scholar
-
Witkowski, M., Weeks, T. L. & Hazen, S. L. Gut microbiota and cardiovascular disease. Circ. Res. 127, 553–570 (2020).
Google Scholar
-
Senthong, V. et al. Plasma trimethylamine N-oxide, a gut microbe-generated phosphatidylcholine metabolite, is associated with atherosclerotic burden. J. Am. Coll. Cardiol. 67, 2620–2628 (2016).
Google Scholar
-
Tang, W. H. et al. Prognostic value of elevated levels of intestinal microbe-generated metabolite trimethylamine-N-oxide in patients with heart failure: refining the gut hypothesis. J. Am. Coll. Cardiol. 64, 1908–1914 (2014).
Google Scholar
-
Tang, W. H. et al. Intestinal microbiota-dependent phosphatidylcholine metabolites, diastolic dysfunction, and adverse clinical outcomes in chronic systolic heart failure. J. Card. Fail 21, 91–96 (2015).
Google Scholar
-
Koeth, R. A. et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 19, 576–585 (2013).
Google Scholar
-
Chen, M. L. et al. Resveratrol attenuates trimethylamine-N-oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut microbiota. mBio 7, e02210–02215 (2016).
Google Scholar
-
Liang, X. et al. Reduction of intestinal trimethylamine by probiotics ameliorated lipid metabolic disorders associated with atherosclerosis. Nutrition 79–80, 110941 (2020).
Google Scholar
-
Valdés-Varela, L., Hernández-Barranco, A. M., Ruas-Madiedo, P. & Gueimonde, M. Effect of Bifidobacterium upon Clostridium difficile growth and toxicity when co-cultured in different prebiotic substrates. Front. Microbiol. https://doi.org/10.3389/fmicb.2016.00738 (2016).
-
Din, A. U. et al. Amelioration of TMAO through probiotics and its potential role in atherosclerosis. Appl. Microbiol. Biotechnol. 103, 9217–9228 (2019).
Google Scholar
-
Hu, T. et al. Short-chain fatty acid metabolism and multiple effects on cardiovascular diseases. Ageing Res. Rev. 81, 101706 (2022).
Google Scholar
-
Hsu, C.-N. et al. Maternal 3,3-dimethyl-1-butanol therapy protects adult male rat offspring against hypertension programmed by perinatal TCDD exposure. Nutrients 13, 3041 (2021).
Google Scholar
-
Turnbaugh, P. J. et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027–1031 (2006).
Google Scholar
-
Tian, Q. et al. Butyrate protects endothelial function through PPARδ/miR-181b signaling. Pharmacol. Res. 169, 105681 (2021).
Google Scholar
-
Yu, Z. et al. Oral supplementation with butyrate improves myocardial ischemia/reperfusion injury via a gut-brain neural circuit. Front. Cardiovasc. Med. 8, 718674 (2021).
Google Scholar
-
Jiang, X., Huang, X., Tong, Y. & Gao, H. Butyrate improves cardiac function and sympathetic neural remodeling following myocardial infarction in rats. Can. J. Physiol. Pharmacol. 98, 391–399 (2020).
Google Scholar
-
Martin-Gallausiaux, C., Marinelli, L., Blottière, H. M., Larraufie, P. & Lapaque, N. SCFA: mechanisms and functional importance in the gut. Proc. Nutr. Soc. 80, 37–49 (2021).
Google Scholar
-
Bourlioux, P., Koletzko, B., Guarner, F. & Braesco, V. The intestine and its microflora are partners for the protection of the host: report on the Danone Symposium “The Intelligent Intestine,” held in Paris, June 14, 2002. Am. J. Clin. Nutr. 78, 675–683 (2003).
Google Scholar
-
Kim, H., Jeong, Y., Kang, S., You, H. J. & Ji, G. E. Co-culture with Bifidobacterium catenulatum improves the growth, gut colonization, and butyrate production of Faecalibacterium prausnitzii: in vitro and in vivo studies. Microorganisms 8 https://doi.org/10.3390/microorganisms8050788 (2020).
-
Bunesova, V., Lacroix, C. & Schwab, C. Mucin cross-feeding of infant Bifidobacteria and Eubacterium hallii. Micro. Ecol. 75, 228–238 (2018).
Google Scholar
-
Moens, F., Weckx, S. & De Vuyst, L. Bifidobacterial inulin-type fructan degradation capacity determines cross-feeding interactions between bifidobacteria and Faecalibacterium prausnitzii. Int. J. Food Microbiol. 231, 76–85 (2016).
Google Scholar
-
Powell-Wiley, T. M. et al. Obesity and cardiovascular disease: a scientific statement from the American Heart Association. Circulation 143, e984–e1010 (2021).
Google Scholar
-
Ndumele, C. E. et al. Obesity and subtypes of incident cardiovascular disease. J. Am. Heart Assoc. 5 https://doi.org/10.1161/jaha.116.003921 (2016).
-
Schellekens, H. et al. Bifidobacterium longum counters the effects of obesity: partial successful translation from rodent to human. EBioMedicine 63, 103176 (2021).
Google Scholar
-
Bagarolli, R. A. et al. Probiotics modulate gut microbiota and improve insulin sensitivity in DIO mice. J. Nutr. Biochem. 50, 16–25 (2017).
Google Scholar
-
Myers, M. G. Jr., Leibel, R. L., Seeley, R. J. & Schwartz, M. W. Obesity and leptin resistance: distinguishing cause from effect. Trends Endocrinol. Metab. 21, 643–651 (2010).
Google Scholar
-
Perakakis, N., Farr, O. M. & Mantzoros, C. S. Leptin in leanness and obesity: JACC state-of-the-art review. J. Am. Coll. Cardiol. 77, 745–760 (2021).
Google Scholar
-
Gabriel, F. C. & Fantuzzi, G. The association of short-chain fatty acids and leptin metabolism: a systematic review. Nutr. Res. 72, 18–35 (2019).
Google Scholar
-
Kojima, M. et al. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 402, 656–660 (1999).
Google Scholar
-
Hosoda, H., Kojima, M. & Kangawa, K. Ghrelin and the regulation of food intake and energy balance. Mol. Inter. 2, 494–503 (2002).
Google Scholar
-
Perelló, M. & Zigman, J. M. The role of ghrelin in reward-based eating. Biol. Psychiatry 72, 347–353 (2012).
Google Scholar
-
English, P. J., Ghatei, M. A., Malik, I. A., Bloom, S. R. & Wilding, J. P. Food fails to suppress ghrelin levels in obese humans. J. Clin. Endocrinol. Metab. 87, 2984 (2002).
Google Scholar
-
Queipo-Ortuño, M. I. et al. Gut microbiota composition in male rat models under different nutritional status and physical activity and its association with serum leptin and ghrelin levels. PLoS ONE 8, e65465 (2013).
Google Scholar
-
Neyrinck, A. M., Schüppel, V. L., Lockett, T., Haller, D. & Delzenne, N. M. Microbiome and metabolic disorders related to obesity: which lessons to learn from experimental models? Trends Food Sci. Technol. 57, 256–264 (2016).
Google Scholar
-
Tang, C., Kong, L., Shan, M., Lu, Z. & Lu, Y. Protective and ameliorating effects of probiotics against diet-induced obesity: a review. Food Res. Int. 147, 110490 (2021).
Google Scholar
-
Salazar, N. et al. Functional effects of EPS-producing bifidobacterium administration on energy metabolic alterations of diet-induced obese mice. Front. Microbiol. 10, 1809 (2019).
Google Scholar
-
Wu, Q., Wang, Q., Fu, J. & Ren, R. Polysaccharides derived from natural sources regulate triglyceride and cholesterol metabolism: a review of the mechanisms. Food Funct. 10, 2330–2339 (2019).
Google Scholar
-
Bigliardi, B. & Galati, F. Innovation trends in the food industry: the case of functional foods. Trends Food Sci. Technol. 31, 118–129 (2013).
Google Scholar
-
Ouali, F., Djouadi, F., Merlet-Bénichou, C., Riveau, B. & Bastin, J. Regulation of fatty acid transport protein and mitochondrial and peroxisomal beta-oxidation gene expression by fatty acids in developing rats. Pediatr. Res. 48, 691–696 (2000).
Google Scholar
-
Canfora, E. E., Meex, R. C. R., Venema, K. & Blaak, E. E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 15, 261–273 (2019).
Google Scholar
-
Machado, A. S. et al. Oral probiotic Bifidobacterium longum supplementation improves metabolic parameters and alters the expression of the renin-angiotensin system in obese mice liver. Biol. Res. Nurs. 23, 100–108 (2021).
Google Scholar
-
Kikuchi, K., Ben Othman, M. & Sakamoto, K. Sterilized bifidobacteria suppressed fat accumulation and blood glucose level. Biochem. Biophys. Res. Commun. 501, 1041–1047 (2018).
Google Scholar
-
Cano, P. G., Santacruz, A., Trejo, F. M. & Sanz, Y. Bifidobacterium CECT 7765 improves metabolic and immunological alterations associated with obesity in high-fat diet-fed mice. Obesity 21, 2310–2321 (2013).
Google Scholar
-
Sternisha, S. M. & Miller, B. G. Molecular and cellular regulation of human glucokinase. Arch. Biochem. Biophys. 663, 199–213 (2019).
Google Scholar
-
Jiang, T. et al. Bifidobacterium longum 070103 fermented milk improve glucose and lipid metabolism disorders by regulating gut microbiota in mice. Nutrients 14 https://doi.org/10.3390/nu14194050 (2022).
-
Nie, Y. & Luo, F. Dietary fiber: an opportunity for a global control of hyperlipidemia. Oxid. Med. Cell Longev. 2021, 5542342 (2021).
Google Scholar
-
Pedro-Botet, J., Millán Núñez-Cortés, J., Chillarón, J. J., Flores-Le Roux, J. A. & Rius, J. Severity of statin-induced adverse effects on muscle and associated conditions: data from the DAMA study. Expert Opin. Drug Saf. 15, 1583–1587 (2016).
Google Scholar
-
Ooi, L. G. & Liong, M. T. Cholesterol-lowering effects of probiotics and prebiotics: a review of in vivo and in vitro findings. Int. J. Mol. Sci. 11, 2499–2522 (2010).
Google Scholar
-
Bordoni, A. et al. Cholesterol-lowering probiotics: in vitro selection and in vivo testing of bifidobacteria. Appl. Microbiol. Biotechnol. 97, 8273–8281 (2013).
Google Scholar
-
Al-Sheraji, S. H. et al. Hypocholesterolaemic effect of yoghurt containing Bifidobacterium pseudocatenulatum G4 or Bifidobacterium longum BB536. Food Chem. 135, 356–361 (2012).
Google Scholar
-
Reynier, M. O. et al. Comparative effects of cholic, chenodeoxycholic, and ursodeoxycholic acids on micellar solubilization and intestinal absorption of cholesterol. J. Lipid Res. 22, 467–473 (1981).
Google Scholar
-
Zanotti, I. et al. Evidence for cholesterol-lowering activity by Bifidobacterium bifidum PRL2010 through gut microbiota modulation. Appl. Microbiol. Biotechnol. 99, 6813–6829 (2015).
Google Scholar
-
Study Group, European Atherosclerosis Society. The recognition and management of hyperlipidaemia in adults: a policy statement of the European Atherosclerosis Society. Eur. Heart J. 9, 571–600 (1988).
-
Nordestgaard, B. G. & Varbo, A. Triglycerides and cardiovascular disease. Lancet 384, 626–635 (2014).
Google Scholar
-
Shaikh, M. et al. Quantitative studies of transfer in vivo of low density, Sf 12-60, and Sf 60-400 lipoproteins between plasma and arterial intima in humans. Arterioscler Thromb. 11, 569–577 (1991).
Google Scholar
-
Nordestgaard, B. G., Tybjaerg-Hansen, A. & Lewis, B. Influx in vivo of low density, intermediate density, and very low density lipoproteins into aortic intimas of genetically hyperlipidemic rabbits. Roles of plasma concentrations, extent of aortic lesion, and lipoprotein particle size as determinants. Arterioscler Thromb. 12, 6–18 (1992).
Google Scholar
-
Nordestgaard, B. G., Stender, S. & Kjeldsen, K. Reduced atherogenesis in cholesterol-fed diabetic rabbits. Giant lipoproteins do not enter the arterial wall. Arteriosclerosis 8, 421–428 (1988).
Google Scholar
-
Goldberg, I. J., Eckel, R. H. & McPherson, R. Triglycerides and heart disease: still a hypothesis. Arterioscler. Thromb. Vasc. Biol. 31, 1716–1725 (2011).
Google Scholar
-
Guardamagna, O. et al. Bifidobacteria supplementation: effects on plasma lipid profiles in dyslipidemic children. Nutrition 30, 831–836 (2014).
Google Scholar
-
Xiao, J. Z. et al. Effects of milk products fermented by Bifidobacterium longum on blood lipids in rats and healthy adult male volunteers. J. Dairy Sci. 86, 2452–2461 (2003).
Google Scholar
-
Fox, C. S. et al. Update on prevention of cardiovascular disease in adults with type 2 diabetes mellitus in light of recent evidence: a scientific statement from the American Heart Association and the American Diabetes Association. Circulation 132, 691–718 (2015).
Google Scholar
-
Benjamin, E. J. et al. Heart disease and stroke statistics-2018 update: a report from the American Heart Association. Circulation 137, e67–e492 (2018).
Google Scholar
-
Ferrannini, E. & Cushman, W. C. Diabetes and hypertension: the bad companions. Lancet 380, 601–610 (2012).
Google Scholar
-
Khan, H. et al. Fasting plasma glucose and incident heart failure risk: a population-based cohort study and new meta-analysis. J. Card. Fail 20, 584–592 (2014).
Google Scholar
-
Kim, S. H. et al. The anti-diabetic activity of Bifidobacterium lactis HY8101 in vitro and in vivo. J. Appl. Microbiol. 117, 834–845 (2014).
Google Scholar
-
Qian, X. et al. Bifidobacterium adolescentis is effective in relieving type 2 diabetes and may be related to its dominant core genome and gut microbiota modulation capacity. Nutrients 14 https://doi.org/10.3390/nu14122479 (2022).
-
Lontchi-Yimagou, E., Sobngwi, E., Matsha, T. E. & Kengne, A. P. Diabetes mellitus and inflammation. Curr. Diab. Rep. 13, 435–444 (2013).
Google Scholar
-
Aoki, R. et al. A proliferative probiotic Bifidobacterium strain in the gut ameliorates progression of metabolic disorders via microbiota modulation and acetate elevation. Sci. Rep. 7, 43522 (2017).
Google Scholar
-
Arnett, D. K. et al. 2019 ACC/AHA guideline on the primary prevention of cardiovascular disease: a report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 140, e596–e646 (2019).
-
Fuchs, F. D. & Whelton, P. K. High blood pressure and cardiovascular disease. Hypertension 75, 285–292 (2020).
Google Scholar
-
Romão da Silva, L. F. et al. Effects of probiotic therapy on cardio-metabolic parameters and autonomic modulation in hypertensive women: a randomized, triple-blind, placebo-controlled trial. Food Funct. 11, 7152–7163 (2020).
Google Scholar
-
Mähler, A., Wilck, N., Rauch, G., Dechend, R. & Müller, D. N. Effect of a probiotic on blood pressure in grade 1 hypertension (HYPRO): protocol of a randomized controlled study. Trials 21, 1032 (2020).
-
Robles-Vera, I. et al. Probiotic Bifidobacterium breve prevents DOCA-salt hypertension. FASEB J. 34, 13626–13640 (2020).
Google Scholar
-
Robles-Vera, I. et al. Probiotics prevent dysbiosis and the rise in blood pressure in genetic hypertension: role of short-chain fatty acids. Mol. Nutr. Food Res. 64, e1900616 (2020).
Google Scholar
-
Rafieian-Kopaei, M., Setorki, M., Doudi, M., Baradaran, A. & Nasri, H. Atherosclerosis: process, indicators, risk factors and new hopes. Int. J. Prev. Med. 5, 927–946 (2014).
-
Hansson, G. K. & Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 12, 204–212 (2011).
Google Scholar
-
Abdi, M., Esmaeili Gouvarchin Ghaleh, H. & Ranjbar, R. Lactobacilli and Bifidobacterium as anti-atherosclerotic agents. Iran. J. Basic Med. Sci. 25, 934–946 (2022).
-
Ataie-Jafari, A., Larijani, B., Alavi Majd, H. & Tahbaz, F. Cholesterol-lowering effect of probiotic yogurt in comparison with ordinary yogurt in mildly to moderately hypercholesterolemic subjects. Ann. Nutr. Metab. 54, 22–27 (2009).
Google Scholar
-
Chiu, H. F., Fang, C. Y., Shen, Y. C., Venkatakrishnan, K. & Wang, C. K. Efficacy of probiotic milk formula on blood lipid and intestinal function in mild hypercholesterolemic volunteers: a placebo-control, randomized clinical trial. Probiotics Antimicrob. Proteins 13, 624–632 (2021).
Google Scholar
-
Lee, Y. et al. Effects of Bifidobacterium animalis subsp. lactis BB-12(®) on the lipid/lipoprotein profile and short chain fatty acids in healthy young adults: a randomized controlled trial. Nutr. J. 16, 39 (2017).
Google Scholar
-
Ejtahed, H. S. et al. Effect of probiotic yogurt containing Lactobacillus acidophilus and Bifidobacterium lactis on lipid profile in individuals with type 2 diabetes mellitus. J. Dairy Sci. 94, 3288–3294 (2011).
Google Scholar
-
Mazloom, Z., Yousefinejad, A. & Dabbaghmanesh, M. H. Effect of probiotics on lipid profile, glycemic control, insulin action, oxidative stress, and inflammatory markers in patients with type 2 diabetes: a clinical trial. Iran. J. Med. Sci. 38, 38–43 (2013).
-
Chen, S. et al. Effects of probiotic supplementation on serum trimethylamine-N-oxide level and gut microbiota composition in young males: a double-blinded randomized controlled trial. Eur. J. Nutr. 60, 747–758 (2021).
Google Scholar
-
Matsumoto, M., Kitada, Y., Shimomura, Y. & Naito, Y. Bifidobacterium animalis subsp. lactis LKM512 reduces levels of intestinal trimethylamine produced by intestinal microbiota in healthy volunteers: a double-blind, placebo-controlled study. J. Funct. Foods 36, 94–101 (2017).
Google Scholar
-
Brunt, V. E. et al. Trimethylamine-N-oxide promotes age-related vascular oxidative stress and endothelial dysfunction in mice and healthy humans. Hypertension 76, 101–112 (2020).
Google Scholar
-
Wang, Z. et al. Measurement of trimethylamine-N-oxide by stable isotope dilution liquid chromatography tandem mass spectrometry. Anal. Biochem. 455, 35–40 (2014).
Google Scholar
-
Pierce, G. L., Roy, S. J. & Gimblet, C. J. The gut-arterial stiffness axis: is TMAO a novel target to prevent age-related aortic stiffening? Hypertension 78, 512–515 (2021).
Google Scholar
-
van den Munckhof, I. C. L. et al. Role of gut microbiota in chronic low-grade inflammation as potential driver for atherosclerotic cardiovascular disease: a systematic review of human studies. Obes. Rev. 19, 1719–1734 (2018).
Google Scholar
-
Raygan, F. et al. The effects of probiotic supplementation on metabolic status in type 2 diabetic patients with coronary heart disease. Diabetol. Metab. Syndr. 10, 51 (2018).
Google Scholar
-
Farrokhian, A. et al. The effects of synbiotic supplementation on carotid intima-media thickness, biomarkers of inflammation, and oxidative stress in people with overweight, diabetes, and coronary heart disease: a randomized, double-blind, placebo-controlled trial. Probiotics Antimicrob. Proteins 11, 133–142 (2019).
Google Scholar
-
Szulińska, M. et al. Multispecies probiotic supplementation favorably affects vascular function and reduces arterial stiffness in obese postmenopausal women—a 12-week placebo-controlled and randomized clinical study. Nutrients 10 https://doi.org/10.3390/nu10111672 (2018).
-
Borshchev, Y. Y. et al. Effects of tetracycline on myocardial infarct size in obese rats with chemically-induced colitis. PLoS ONE 14, e0225185 (2019).
Google Scholar
-
Kwun, J. S. et al. Comparison of thrombus, gut, and oral microbiomes in Korean patients with ST-elevation myocardial infarction: a case-control study. Exp. Mol. Med. 52, 2069–2079 (2020).
Google Scholar
-
Lam, V. et al. Intestinal microbiota determine severity of myocardial infarction in rats. FASEB J. 26, 1727–1735 (2012).
Google Scholar
-
Danilo, C. A. et al. Bifidobacterium animalis subsp. lactis 420 mitigates the pathological impact of myocardial infarction in the mouse. Benef. Microbes 8, 257–269 (2017).
Google Scholar
-
Sadeghzadeh, J. et al. The effect of oral consumption of probiotics in prevention of heart injury in a rat myocardial infarction model: a histopathological, hemodynamic and biochemical evaluation. Iran. Biomed. J. 21, 174–181 (2017).
Google Scholar
-
Frasure-Smith, N., Lespérance, F. & Talajic, M. Depression following myocardial infarction: impact on 6-month survival. Jama 270, 1819–1825 (1993).
Google Scholar
-
Arseneault-Bréard, J. et al. Combination of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 reduces post-myocardial infarction depression symptoms and restores intestinal permeability in a rat model. Br. J. Nutr. 107, 1793–1799 (2012).
Google Scholar
-
Gilbert, K. et al. Attenuation of post-myocardial infarction depression in rats by n-3 fatty acids or probiotics starting after the onset of reperfusion. Br. J. Nutr. 109, 50–56 (2013).
Google Scholar
-
Trudeau, F. et al. Bifidobacterium longum R0175 attenuates post-myocardial infarction depressive-like behaviour in rats. PLoS ONE 14, e0215101 (2019).
Google Scholar
-
Girard, S. A. et al. Lactobacillus helveticus and Bifidobacterium longum taken in combination reduce the apoptosis propensity in the limbic system after myocardial infarction in a rat model. Br. J. Nutr. 102, 1420–1425 (2009).
Google Scholar
-
Moludi, J., Alizadeh, M., Mohammadzad, M. H. S. & Davari, M. The effect of probiotic supplementation on depressive symptoms and quality of life in patients after myocardial infarction: results of a preliminary double-blind clinical trial. Psychosom. Med. 81, 770–777 (2019).
Google Scholar
-
Yancy, C. W. et al. 2017 ACC/AHA/HFSA focused update of the 2013 ACCF/AHA guideline for the management of heart failure: a report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines and the Heart Failure Society of America. Circulation 136, e137–e161 (2017).
Google Scholar
-
Messerli, F. H., Rimoldi, S. F. & Bangalore, S. The transition from hypertension to heart failure: contemporary update. JACC Heart Fail 5, 543–551 (2017).
Google Scholar
-
Flaherty, J. D. et al. Acute heart failure syndromes in patients with coronary artery disease early assessment and treatment. J. Am. Coll. Cardiol. 53, 254–263 (2009).
Google Scholar
-
Jenča, D. et al. Heart failure after myocardial infarction: incidence and predictors. ESC Heart Fail 8, 222–237 (2021).
Google Scholar
-
McMurray, J. J. et al. ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: The Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure 2012 of the European Society of Cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 33, 1787–1847 (2012).
Google Scholar
-
Taylor, C. J. et al. Trends in survival after a diagnosis of heart failure in the United Kingdom 2000-2017: population based cohort study. BMJ 364, l223 (2019).
Google Scholar
-
Chorianopoulos, E. et al. Decreased soluble TWEAK levels predict an adverse prognosis in patients with chronic stable heart failure. Eur. J. Heart Fail 11, 1050–1056 (2009).
Google Scholar
-
Richter, B. et al. Differences in the predictive value of tumor necrosis factor-like weak inducer of apoptosis (TWEAK) in advanced ischemic and non-ischemic heart failure. Atherosclerosis 213, 545–548 (2010).
Google Scholar
-
Blanco-Colio, L. M. et al. Vascular proteomics and the discovery process of clinical biomarkers: the case of TWEAK. Proteom. Clin. Appl. 5, 281–288 (2011).
Google Scholar
-
Pourrajab, B. et al. The impact of probiotic yogurt versus ordinary yogurt on serum sTWEAK, sCD163, ADMA, LCAT and BUN in patients with chronic heart failure: a randomized, triple-blind, controlled trial. J. Sci. Food Agric. 102, 6024–6035 (2022).
Google Scholar
-
Pourrajab, B. et al. Comparison of probiotic yogurt and ordinary yogurt consumption on serum Pentraxin3, NT-proBNP, oxLDL, and ApoB100 in patients with chronic heart failure: a randomized, triple-blind, controlled trial. Food Funct. 11, 10000–10010 (2020).
Google Scholar
-
Yu, H. et al. Intervention effect of probiotics in gastric cancer patients with complications of coronary heart disease and heart failure. J. Oncol. 2021, 1620891 (2021).
Google Scholar
-
Pan, M. et al. Genomic and epigenetic landscapes drive CRISPR-based genome editing in Bifidobacterium. Proc. Natl Acad. Sci. USA 119, e2205068119 (2022).
Google Scholar
-
O’Connell Motherway, M. et al. Characterization of ApuB, an extracellular type II amylopullulanase from Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 74, 6271–6279 (2008).
Google Scholar
-
Sakanaka, M. et al. A transposon mutagenesis system for Bifidobacterium longum subsp. longum based on an IS3 family insertion sequence, ISBlo11. Appl. Environ. Microbiol. 84 https://doi.org/10.1128/aem.00824-18 (2018).
-
Zuo, F., Zeng, Z., Hammarström, L. & Marcotte, H. Inducible plasmid self-destruction (IPSD) assisted genome engineering in Lactobacilli and Bifidobacteria. ACS Synth. Biol. 8, 1723–1729 (2019).
Google Scholar
-
Zuo, F. & Marcotte, H. Advancing mechanistic understanding and bioengineering of probiotic lactobacilli and bifidobacteria by genome editing. Curr. Opin. Biotechnol. 70, 75–82 (2021).
Google Scholar
-
O’Connell Motherway, M. et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl Acad. Sci. USA 108, 11217–11222 (2011).
Google Scholar
-
Zuo, F., Chen, S. & Marcotte, H. Engineer probiotic bifidobacteria for food and biomedical applications – current status and future prospective. Biotechnol. Adv. 45, 107654 (2020).
Google Scholar
Acknowledgements
This study was supported by the Cooperation Projectors of Chunhui Plan of the Ministry of Education of China (No. HZKY20220563), the Central Government Guides the Local Science and Technology Development Special Fund (No. 2022ZYD0084), the Key R&D projects (No. 2022YFS0607, 2022YFS0635, 2022YFS0627), the Youth Science and Technology Innovation Research Team of the Science and Technology Department of Sichuan province of China (No. 2021JDTD0008), the Open Research Fund of Chengdu University of Traditional Chinese Medicine State Key Laboratory of Southwestern Chinese Medicine Resources (No. SKLTCM2022027), Key Research and Development Projectors of Luzhou (No. 2021-SYF-26, 2022-SYF-85), the cooperation Project (No. 230007-01SZ, 230008-01SZ) of Sichuan Credit Pharmaceutical CO., Ltd., Central Nervous System Drug Key Laboratory of Sichuan Province, Chongqing Traditional Chinese Medicine Inheritance and Innovation Team Construction Project “Traditional Chinese Medicine New Drug and Safety Research Inheritance and Innovation Team” (No. 2022-8), Key Research and Development Projectors (No. 2022CDLZ-10) of Sichuan University-Luzhou, the Innovation Team and Talents Cultivation Program (No. ZYYCXTD-C-202207) of National Administration of Traditional Chinese Medicine.
Author information
Authors and Affiliations
Contributions
J.T., Y.W., and C.P.: Writing—original draft preparation; L.Z.: Writing—review & editing; N.Z. and C.P.: Project administration; P.S., J.C., and L.X.: Conceptualization; W.Z. and Y.Z.: Supervision; H.L., T.C., Q.Z., and S.Y.: Acquisition of funds; W.R.: Visualization, investigation.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Reprints and Permissions
About this article
Cite this article
Tang, J., Wei, Y., Pi, C. et al. The therapeutic value of bifidobacteria in cardiovascular disease.
npj Biofilms Microbiomes 9, 82 (2023). https://doi.org/10.1038/s41522-023-00448-7
-
Received: 04 June 2023
-
Accepted: 03 October 2023
-
Published: 30 October 2023
-
DOI: https://doi.org/10.1038/s41522-023-00448-7